种群生态学
种群生态学是生态学的核心分支之一,主要研究同一物种的个体在一定时间和空间范围内所形成的集合体——即“种群”的结构、功能与动态变化。它不仅关注种群数量的增减,还涉及到种群的年龄结构、性别比例、分布格局及其与环境因素之间的关系。理解种群的动态变化规律,对于生物多样性保护、野生动物管理、渔业和林业资源可持续利用以及农业害虫防治等领域都具有不可替代的科学与实际指导意义。例如,通过掌握某些濒危动物的种群变化趋势,可以制定更有效的保护措施;了解农业害虫种群的爆发规律,能够为绿色植保提供理论依据。
在我们的日常生活中,种群生态学的原理同样随处可见。春天成群归来的候鸟,揭示了迁徙种群对环境变化的响应能力;田间害虫数量的骤然增加,反映了极端天气或生物天敌减少对种群波动的影响;城市里流浪猫数量的长期变化,则提示着人类活动和自然环境共同塑造了城市动物种群的命运。这些现象既展示了种群生态学的研究对象,也显示了其在实际生活中的重要作用。无论是科学研究还是生态管理,种群生态学都为我们理解和调控自然界中生命的数量与结构,提供了坚实的理论基础。
种群的基本特征与参数
种群是生态学研究的基本单位之一,它指的是在一定时间和空间范围内,同一物种的所有个体组成的集合。要准确描述一个种群的状态,我们需要掌握几个核心参数。
种群密度是最直观的种群特征,它表示单位面积或单位体积内的个体数量。以长江流域的扬子鳄为例,生态学家通过多年监测发现,野生扬子鳄的种群密度在上世纪90年代仅为每平方公里0.5条左右,这一极低的密度反映了该物种濒危的严峻现实。而在人工保护区域,通过科学管理,密度可以提升到每平方公里5-8条,种群恢复显著。
种群的出生率和死亡率是决定种群数量变化的两个关键因素。出生率指单位时间内新增个体数占种群总数的比例,死亡率则指单位时间内死亡个体数占总数的比例。在四川卧龙自然保护区的大熊猫种群研究中,科学家发现,成年雌性大熊猫的年均繁殖率约为0.3-0.4,也就是说每只成年雌性平均每年生育0.3-0.4只幼崽。同时,幼年大熊猫的死亡率在野外环境下可达30%-40%,这主要与食物短缺、疾病和天敌捕食有关。
种群的出生率减去死亡率,就得到了种群的自然增长率。当自然增长率为正值时,种群数量增加;为负值时,种群数量减少;为零时,种群数量保持稳定。
年龄结构是种群的重要特征之一,它反映了不同年龄组个体在种群中的比例。通常我们将种群个体分为三个年龄组:幼年前生殖期、成年生殖期和老年后生殖期。不同的年龄结构预示着种群未来的发展趋势。在东北虎豹国家公园的野生东北虎种群中,研究人员通过红外相机监测发现,幼年个体占比从2010年的15%提升到2020年的28%,这种增长型的年龄结构表明,随着生态环境的改善和保护力度的加强,东北虎种群正处于恢复性增长阶段。
性别比例也是影响种群繁殖潜力的重要因素。大多数动物种群的性别比例接近1:1,但在某些特殊情况下会出现偏离。以中国特有的藏羚羊为例,在可可西里地区的研究显示,成年藏羚羊的雌雄比例约为3:1,这是因为雄性个体在争夺配偶的过程中死亡率较高,同时雄性个体还面临着更大的捕猎压力。
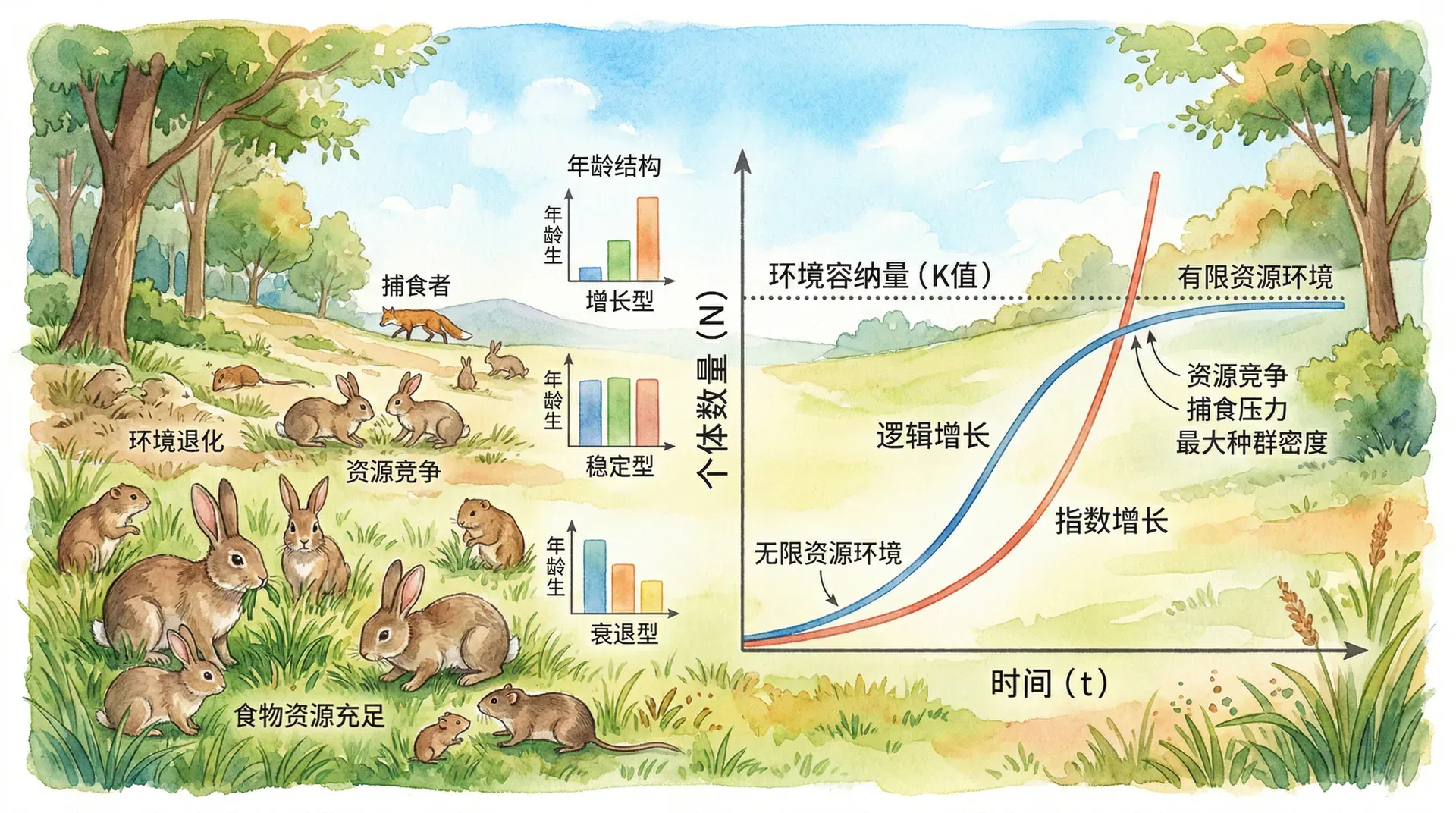
从上图可以看出,增长型种群幼年个体数量最多,预示着种群将快速增长;稳定型种群各年龄组比例相对均衡,种群数量将保持稳定;而衰退型种群老年个体比例过高,种群面临衰退风险。
种群增长模型
指数增长模型
当种群处于理想环境中,资源充足、空间无限、没有天敌和疾病时,种群会以其最大能力进行繁殖,这种增长方式称为指数增长。指数增长的数学表达式为:
其中 表示 时刻的种群数量, 表示初始数量, 为种群的内禀增长率, 为自然对数的底。
在现实世界中,真正的指数增长往往只能在短期内或特定条件下实现。一个典型的例子是20世纪80年代引入澳大利亚的欧洲兔,由于缺乏天敌、气候适宜、食物丰富,仅24只兔子在6年内繁殖到数百万只,给当地生态系统造成了巨大破坏。在中国,草原蝗虫在适宜的气候条件下也会出现短期的指数增长,一只雌性蝗虫可产卵50-100粒,在温暖湿润的环境下,一个月内就能完成一个世代,种群数量呈几何级数增长。
上图展示了在理想条件下,种群如何呈现“J”型增长曲线。曲线初期相对平缓,但随着时间推移,增长速度越来越快,形成了陡峭的上升趋势。
逻辑斯谛增长模型
然而,自然界中没有无限的资源,任何环境都有其承载能力的上限,这个上限被称为环境容纳量(用 表示)。当种群数量接近环境容纳量时,由于资源竞争加剧、生存空间减少,种群的增长速度会逐渐放缓,最终趋于稳定。这种增长模式被称为逻辑斯谛增长,其数学表达式为:
逻辑斯谛增长曲线呈现“S”型,可以分为四个阶段。在起始期,种群数量少,增长缓慢;进入加速期后,随着个体数量增加,增长速度加快;到达转折期时,种群数量达到环境容纳量的一半,此时增长速度最快;最后进入饱和期,种群数量接近环境容纳量,增长趋于停滞。
以青海湖裸鲤的种群恢复为例,从2002年开始实施全面禁捕以来,裸鲤资源量从2002年的2592吨恢复到2020年的10.04万吨。在恢复初期(2002-2008年),由于种群基数小,年均增长率约为8%;进入加速期(2008-2015年),年均增长率提升至15%;而在2015年之后,随着种群数量接近青海湖的环境容纳量,增长速度开始放缓,年均增长率降至6%左右。
从图中可以清楚地看到,种群数量在接近环境容纳量时增长趋缓,最终稳定在K值附近波动。在种群数量达到K/2时,也就是环境容纳量的一半时,种群的增长速度达到最大值,这一点在资源管理和物种保护中具有重要的实践意义。
在野生动物管理和渔业资源开发中,通常将种群数量维持在K/2左右,此时种群既有较快的恢复能力,又能持续提供一定的可利用资源。
种群调节机制与环境容纳量
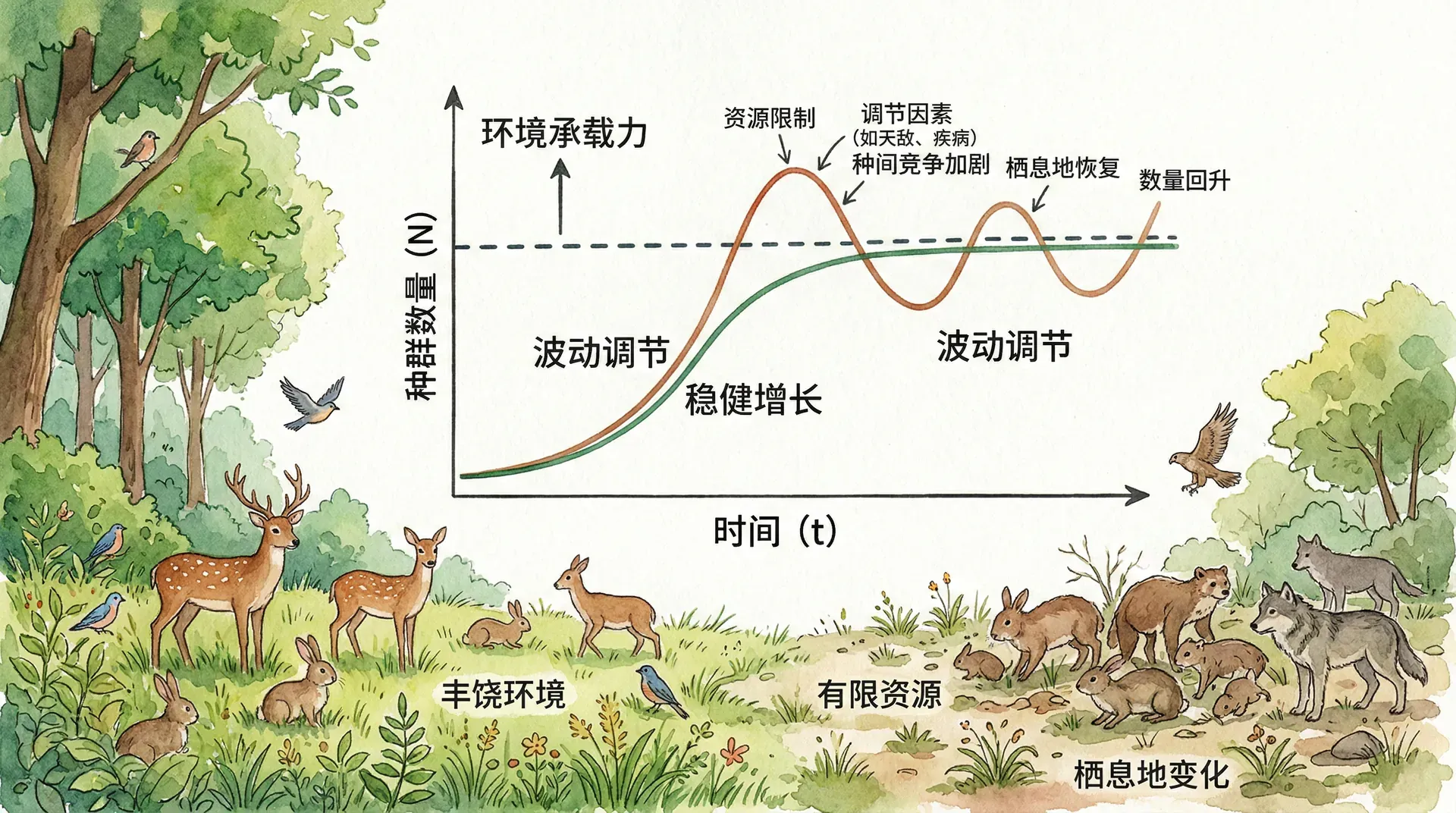
种群数量不会无限制地增长,自然界中存在多种调节机制来限制种群规模。这些调节机制可以分为密度制约因素和非密度制约因素两大类。
密度制约因素的作用强度随种群密度的变化而变化。当种群密度增加时,个体之间对食物、空间、配偶等资源的竞争加剧,导致出生率下降、死亡率上升。在四川王朗自然保护区对川金丝猴的研究中发现,当猴群规模超过80只时,个体间的社会压力明显增大,表现为攻击行为增多、幼崽存活率下降,最终导致猴群分裂成两个或多个小群体。这种行为调节机制有效地防止了种群过度增长。
近年来,随着天敌数量减少和保护力度加强,东北部分地区野猪种群快速增长。当野猪密度过高时,食物资源变得紧张,野猪不得不扩大活动范围,甚至频繁进入农田觅食,这增加了它们与人类冲突的机会,也提高了疾病传播和意外死亡的风险。当种群密度超过每平方公里8-10头时,疾病暴发的概率显著上升,从而自然调节了种群数量。
非密度制约因素则与种群密度无关,主要包括气候变化、自然灾害等。2008年初的南方雨雪冰冻灾害对许多野生动物种群造成了严重影响,大量鸟类因无法觅食而死亡,这种影响不论种群密度高低都会发生。同样,极端高温或低温、洪涝、干旱等气候事件都可能在短时间内造成种群数量的剧烈波动。
环境容纳量不是一个固定的数值,它会随着环境条件的改变而波动。在内蒙古草原,草食动物的环境容纳量与降雨量密切相关。在降雨充沛的年份,牧草生长旺盛,草原的环境容纳量可能比干旱年份高出30%-50%。因此,合理的草原管理需要根据年际间的降雨变化动态调整放牧强度,而不能简单地按照固定的载畜量来管理。
生命表与存活曲线
生命表最早起源于人口统计学,后来被引入生态学研究中,用于描述种群中个体从出生到死亡的全过程。通过生命表,我们可以清晰地了解不同年龄阶段个体的存活状况和死亡风险。
一个完整的生命表包含多个重要参数。年龄组表示个体所处的生命阶段;存活数表示该年龄组起始时的存活个体数;死亡数表示在该年龄段内死亡的个体数;存活率则是存活数占初始个体数的比例。以野生朱鹮为例,研究人员通过多年追踪发现,朱鹮在第一年的死亡率最高,约为40%,这主要是因为雏鸟容易受到天敌捕食、恶劣天气影响以及食物短缺的威胁。度过第一年后,亚成年朱鹮的年死亡率降至15%左右,而成年朱鹮的年死亡率稳定在10%以内。
根据生命表数据,我们可以绘制存活曲线,它直观地展示了种群个体在不同年龄段的存活情况。生态学家将存活曲线归纳为三种基本类型。
凸型曲线(Ⅰ型)的特点是幼年和成年期存活率很高,只有到了老年期才出现大量死亡。这种类型常见于亲代对后代有较强保护能力的物种,如大型哺乳动物。以亚洲象为例,幼象在母象的精心照顾下,前几年的死亡率很低,只有当进入老年后,由于牙齿磨损、疾病增多,死亡率才急剧上升。
对角线型(Ⅱ型)表示各年龄段的死亡率相对恒定,存活曲线接近直线。许多鸟类和小型哺乳动物属于这一类型。黄腹角雉是中国特有的珍稀鸟类,研究表明,除了繁殖季节外,成年黄腹角雉各年龄段的死亡率相对稳定,每年约为20%左右。
凹型曲线(Ⅲ型)的特征是幼年期死亡率极高,但一旦存活下来进入成年期,就能较长时间地生存。这种类型普遍存在于繁殖力强、亲代对后代投资少的物种中。中国的四大家鱼(青鱼、草鱼、鲢鱼、鳙鱼)就是典型代表,一尾雌鱼可产卵数十万至上百万粒,但受精卵孵化后,大部分鱼苗会被其他鱼类捕食或因环境不适而死亡,真正能长成幼鱼的比例不到1%。然而,一旦长成成鱼,它们的存活时间可以达到10年甚至更长。
通过分析不同物种的存活曲线类型,我们可以制定更有针对性的保护策略。对于凸型曲线的物种,应重点保护成年个体,维持繁殖种群;对于凹型曲线的物种,则应着重改善幼年个体的生存环境,提高早期存活率。
中国典型种群的动态案例研究
大熊猫种群的恢复
大熊猫是中国的国宝,也是世界生物多样性保护的旗舰物种。根据第四次全国大熊猫调查数据,截至2014年,野生大熊猫种群数量为1864只,比2003年增长了16.8%。这一成就离不开科学的种群管理策略。
在四川省,大熊猫种群主要分布在岷山、邛崃山、大相岭、小相岭和凉山五大山系。每个山系的大熊猫种群相对独立,长期的地理隔离导致了遗传多样性下降的风险。为此,生态学家提出了建立大熊猫走廊带的方案,通过恢复连接不同栖息地的森林植被,促进种群间的基因交流。在大熊猫国家公园建立后,这一方案得到了系统实施,多个隔离种群之间的连通性显著改善。
栖息地质量是影响大熊猫种群数量的关键因素。大熊猫的主食竹子具有周期性开花死亡的特性,通常30-60年开花一次。1983年岷山地区箭竹大面积开花枯死,导致约250只大熊猫因食物短缺而死亡,种群数量急剧下降。这一事件促使科学家深入研究竹子的开花周期,在不同海拔高度营造多种竹类混交林,确保当一种竹子开花时,大熊猫还能取食其他种类的竹子。
从图表可以看出,经过40年的保护努力,大熊猫野生种群数量稳步回升,特别是在1999年之后增长明显加快,这与天然林保护工程的实施、自然保护区网络的建立以及社区共管模式的推广密切相关。
长江江豚的种群危机

长江江豚是长江特有的淡水鲸类动物,被誉为长江生态系统健康的指示物种。然而,近几十年来,江豚种群数量持续下降,保护形势十分严峻。
根据长江江豚科学考察数据,2006年长江江豚种群数量约为1800头,2012年下降至1045头,2017年进一步下降至1012头。种群数量的快速下降主要归因于栖息地退化、食物资源减少、航运干扰、水污染以及非法捕捞误捕等多重因素。特别是在长江中下游的部分江段,由于高强度的人类活动,江豚的环境容纳量大幅降低。
为挽救这一濒危物种,中国启动了长江江豚迁地保护工程。在湖北监利何王庙、湖南洞庭湖、安徽安庆西江等地建立了迁地保护区,通过人工捕捞将部分江豚迁移到相对封闭、环境较好的水域中繁育。监测数据显示,何王庙保护区的江豚种群从最初的5头增加到2020年的80余头,表现出良好的繁殖能力。这一成功经验为其他淡水鲸类的保护提供了宝贵的借鉴。
2021年开始实施的长江十年禁渔政策,为江豚种群的恢复创造了历史性机遇。随着渔业活动的停止,长江中的鱼类资源开始恢复,江豚的食物供应得到改善。同时,航运管理的规范化减少了船只对江豚的干扰和伤害。初步监测显示,部分江段的江豚种群数量出现了止跌回升的迹象。
从表格数据可以看出,长江江豚的保护工作正在取得初步成效,种群下降趋势得到遏制,并开始呈现恢复性增长。但要实现种群的长期稳定和增长,仍需持续加强栖息地保护、减少人为干扰、恢复鱼类资源。
内蒙古草原鼠类种群的周期性波动
在内蒙古草原生态系统中,啮齿动物(主要是布氏田鼠、达乌尔黄鼠)的种群数量呈现出明显的周期性波动,一般为3-4年一个周期。这种周期性波动不仅影响草原植被,还与草原鸟类、食肉动物的种群动态密切相关。
当鼠类种群处于低谷期时,每公顷草原的鼠类密度约为20-50只。此时草原植被恢复良好,食物充足,鼠类的繁殖率提高,种群开始增长。进入增长期后,充足的食物供应和较少的种内竞争促使鼠类快速繁殖,每只雌鼠一年可产4-6胎,每胎5-8只幼鼠,种群密度迅速上升。
当种群密度达到高峰期,每公顷可达300-500只甚至更多时,草原植被被严重破坏,鼠类开始争夺有限的食物和领地,种内竞争激烈。同时,高密度的鼠类种群为天敌提供了丰富的食物来源,猛禽如草原雕、纵纹腹小鸮等的数量随之增加,捕食压力加大。此外,高密度还导致疫病暴发,大量鼠类死亡。这些因素共同作用,使得鼠类种群数量急剧下降,进入衰退期。
从图中可以清晰地看到,鼠类密度与植被覆盖度呈现明显的负相关关系。当鼠类密度处于高峰时,植被覆盖度降至最低;而当鼠类密度下降后,植被逐渐恢复。这种周期性波动是草原生态系统自我调节的体现。
虽然鼠类种群的周期性波动是自然现象,但当草原退化严重、天敌减少时,这种调节机制可能失效,导致鼠害持续发生。因此,草原管理需要综合考虑植被恢复、天敌保护、适度放牧等多种措施。
通过这三个案例可以看出,种群动态受到多种因素的综合影响,包括栖息地质量、食物资源、天敌关系、人为干扰等。科学的种群管理需要建立在对这些因素深入理解的基础上,采取系统性的保护措施,才能实现种群的可持续发展。
本节练习
1. 某自然保护区对一种珍稀鸟类进行种群调查,2020年记录到成年个体300只,当年新增幼鸟90只,死亡个体45只。请计算该鸟类种群的出生率、死亡率和自然增长率,并预测如果保持这一增长速度,2025年该种群的数量约为多少只?(假设增长率保持不变)
2. 下表为某湖泊中鲤鱼种群在不同密度下的繁殖数据。请分析说明该种群受到何种调节机制的影响,并解释为什么会出现这种变化趋势。
3. 在一个面积为100平方公里的保护区中,科学家估算某食草动物的环境容纳量K为2000只。目前该种群数量为600只,内禀增长率r为0.2。请计算:
(1) 当前该种群的瞬时增长速率是多少?
(2) 在什么种群规模时,该种群的增长速率达到最大?此时的增长速率是多少?
(3) 从保护生物学角度,说明为什么不应该让种群数量达到环境容纳量K?
4. 草原生态系统中,狼与羊的种群数量常呈现周期性波动,两者的数量变化存在明显的时间滞后关系。请运用种群生态学原理,解释这种周期性波动现象的形成机制,并说明这种波动对维持生态系统稳定性的意义。
5. 近年来,中国部分地区野猪种群数量快速增长,频繁进入农田和居民区,造成农作物损失和人身安全隐患。请从种群生态学角度分析野猪种群快速增长的原因,并提出科学合理的种群调控建议。