器官发生与形态建成
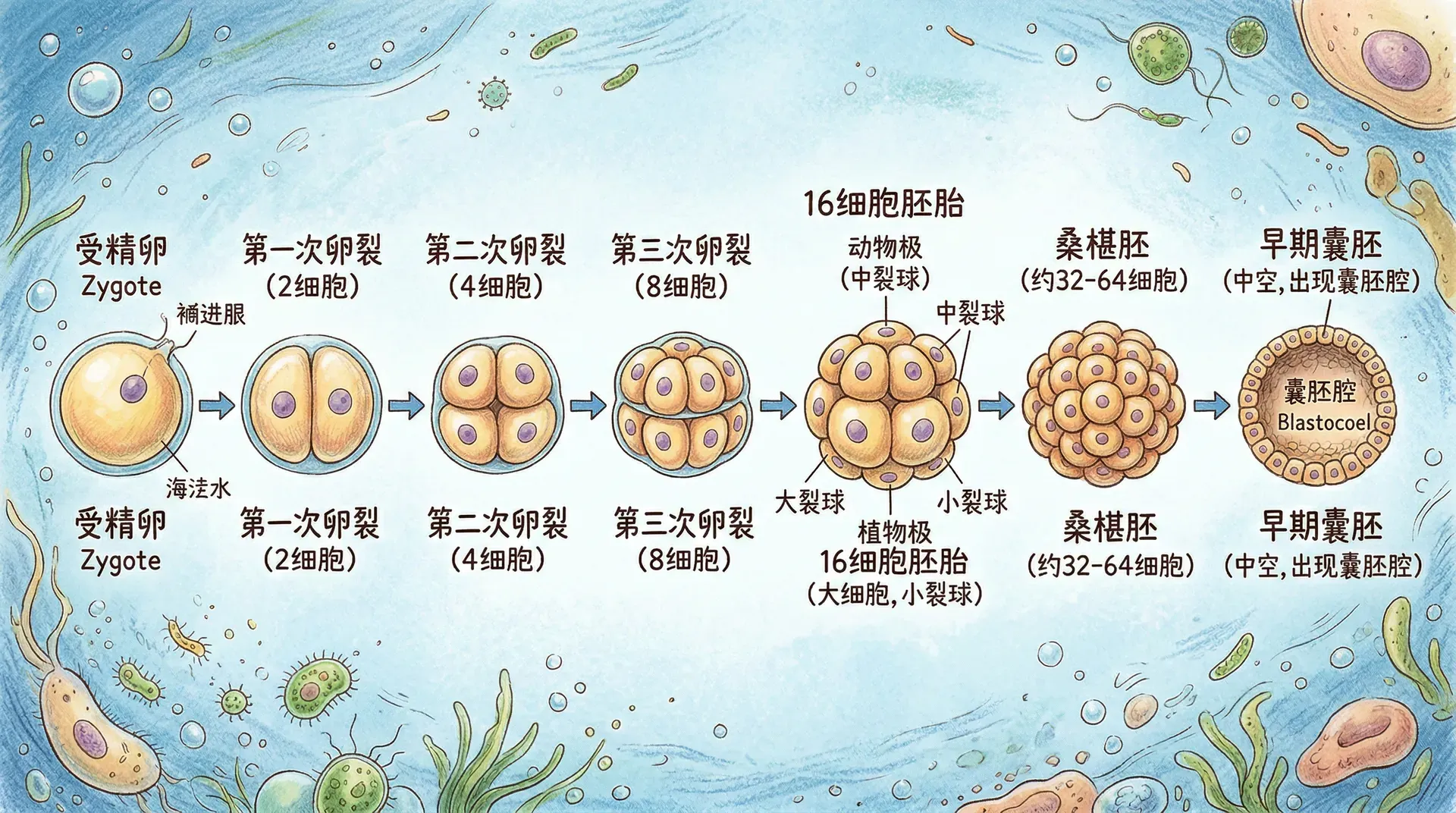
在经典发育生物学中,海胆早期胚胎的分裂和细胞命运决定机制为科学家们提供了许多基础性理论。19世纪的移植实验表明,将一个海胆胚胎的单个细胞分离后,每个细胞都有能力发育成一个完整的幼体,这揭示了早期胚胎细胞具备全能性。此外,蝾螈的胚胎移植实验首次证明了诱导作用的存在,即部分胚胎组织可以影响邻近组织的发育方向,引发重要结构的形成。这些实验为理解多细胞机体的空间格局建立奠定了基础。
近年来,单细胞测序技术和活细胞成像手段推动了发育过程研究的精细化。研究者能够追踪单个细胞在胚胎中的运动路径,分析基因表达的动态变化。例如,在小鼠胚胎中,单细胞RNA测序帮助揭示了肝脏、胰腺等器官原基的细胞组成和谱系关系。有意思的是,对果蝇、斑马鱼等模式生物的研究,推动了调控基因组和信号通路功能的深入解析,为人类疾病机制的研究提供了模型和启示。
神经系统的发育过程
神经系统的发育是胚胎最早启动的器官发生之一。在人类胚胎发育第三周,原肠胚形成后,背侧外胚层细胞在脊索诱导下开始向神经系统方向分化,这一过程称为神经诱导。
神经管的形成
神经管的形成是神经系统发育的起点。在脊索分泌的信号作用下,胚胎背侧外胚层细胞增厚形成神经板,神经板中部下陷形成神经沟,两侧隆起为神经褶。第四周,神经褶向中线靠拢融合,形成封闭的神经管。闭合自躯干向头尾两端推进,最后在两端闭合。
神经管闭合的时机极为重要。受孕后第28天内如果未能及时闭合,会导致诸如无脑畸形或脊柱裂等神经管缺陷。
神经管闭合后,管腔成为脑室和脊髓中央管,管壁逐步分为五大脑区:
神经嵴细胞的迁移与分化
神经褶融合时,神经板与外胚层交界的细胞分离——这些即为神经嵴细胞,拥有强大迁移与多向分化的能力。神经嵴细胞迁移路径及分化产物如下:
例如,在中国南方常见的“白化病”,其发生机制是神经嵴来的黑色素细胞因酶基因突变,不能合成黑色素,导致皮肤、毛发、虹膜变浅。
神经元的产生与分层
神经管壁起初为单层神经上皮,不断分裂以增加细胞数。随发育进展,逐渐形成以下功能性层次:
神经前体可不对称分裂,一部分保持前体属性,另一部分分化为神经母细胞。神经母细胞迁移至套层后分裂、停止,再分化成熟为神经元,最终迁移到边缘层,发射轴突与树突,构建神经网络。
大脑皮层发育突出特点是“由内向外”分层。深层神经元先生成,后续新生神经元越过旧层,定居在更外层,形成六层结构。
如上图所见,胚胎期大脑皮层神经元在发育中期快速增加,脊髓神经元数量则增长较为平稳。这展示了各脑区神经元产生的时序性差异。
心血管系统的形成机制
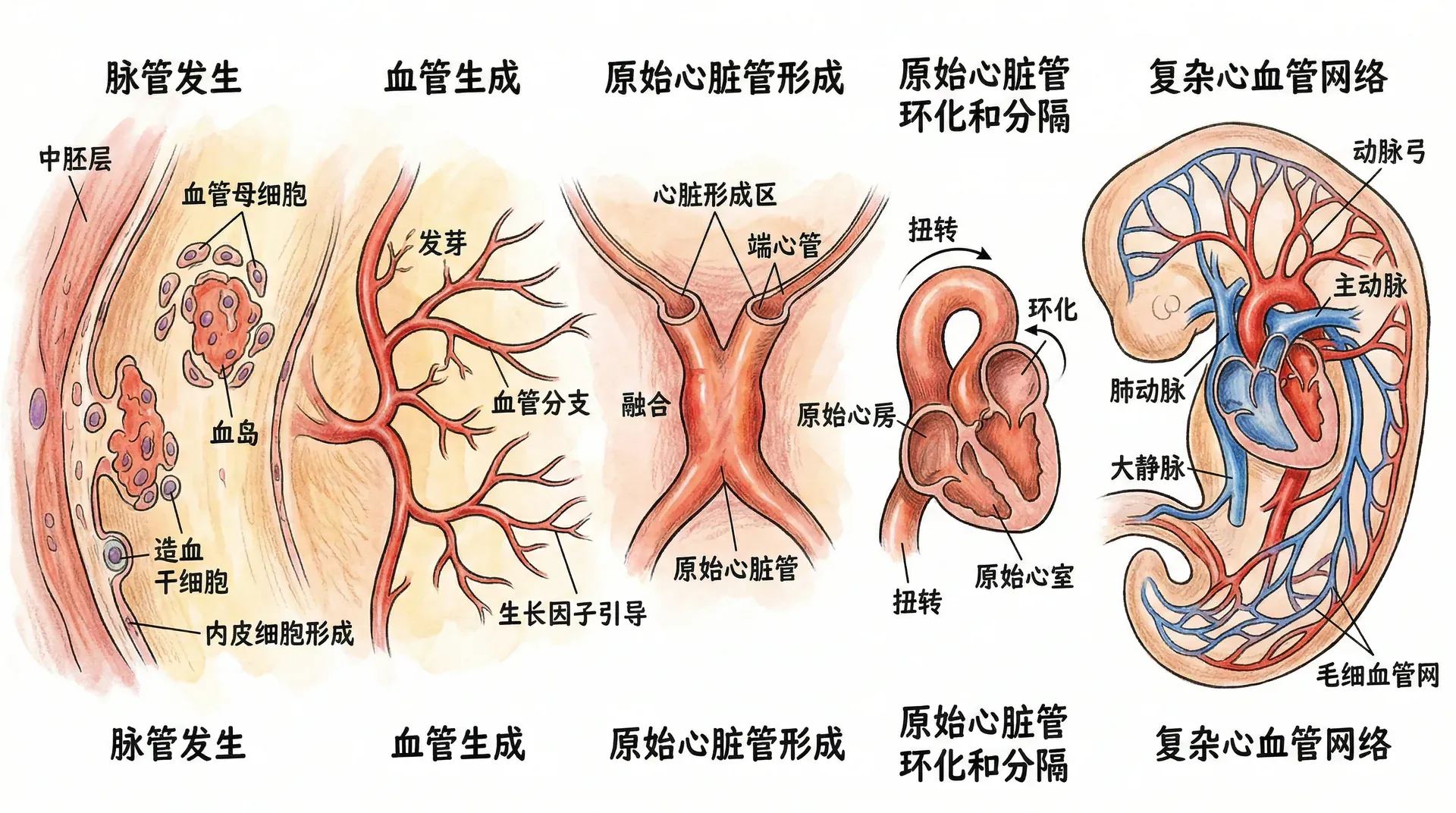
心血管系统在胚胎最早发功能。到第三周末,心脏已能跳动,将血液输送至原始血管网络,实现营养与氧气运输。
心脏的早期发育
心脏起源于头端中胚层,两片弯月形心源区内卷,形成两根心内膜管。随着胚体折叠及头部抬升,心内膜管移至腹侧并融合成单一原始心管。
原始心管自头到尾分区如下:
心管在早期尚未分腔时已具备蠕动性收缩能力,其节律性收缩自静脉窦端起始,波状向头端(动脉端)推进,从而保证血液按照单一方向流动。这一原始心管的推进型收缩机制,是胚胎循环建立的基础,也为后期心脏分化形成完整泵血功能打下了根基。
心脏的弯曲与分隔
第四周,心管扩张快于心包腔,出现C形、S形弯折,心房上移至心室背后,形成心脏左右空间布局。随后心脏分隔发生,形成四腔室结构。主要分隔过程如下:
心脏瓣膜的形成
心瓣膜可分为房室瓣(如二尖、三尖瓣)和动脉瓣(如主动脉、肺动脉瓣),两类形成机制各异:
瓣膜如同心脏的“单向门”,精准地控制血液只能按固定方向流动,有效防止血液回流,保障心腔间的压力分布。健康的瓣膜功能是心脏高效泵血、全身正常供氧的基础,其结构异常会导致循环障碍和临床症状。
血管网络的建立
心脏发育并行,血管在全身形成。两种机制包括:
中国科研团队以斑马鱼为模型,系统研究了VEGF在血管生成的调控作用,为缺血性疾病治疗破题。
上图展示了胚胎心脏在不同发育阶段的心率变化趋势。可以看到,随着胚胎的逐步发育,心率由早期的相对较低水平逐渐升高,反映出心脏结构和功能的快速完善。到第8周左右,心率达到发育过程中的高峰,此时心脏的解剖结构基本成型,能够高效地推动循环。随后,心率略有下降,并保持在一个较为稳定的水平,直至成鱼阶段。这种变化既体现了心脏泵血负荷的动态变化,也反映了循环系统成熟过程中的生理调节机制。
四肢发育与模式形成
四肢的发育为研究空间模式形成机制提供了经典范例。从小小的肢芽,到分明的手指脚趾,细胞通过获取特定位置信息和信号梯度,逐步决定自身的命运、极性与结构。
肢芽的启动与生长
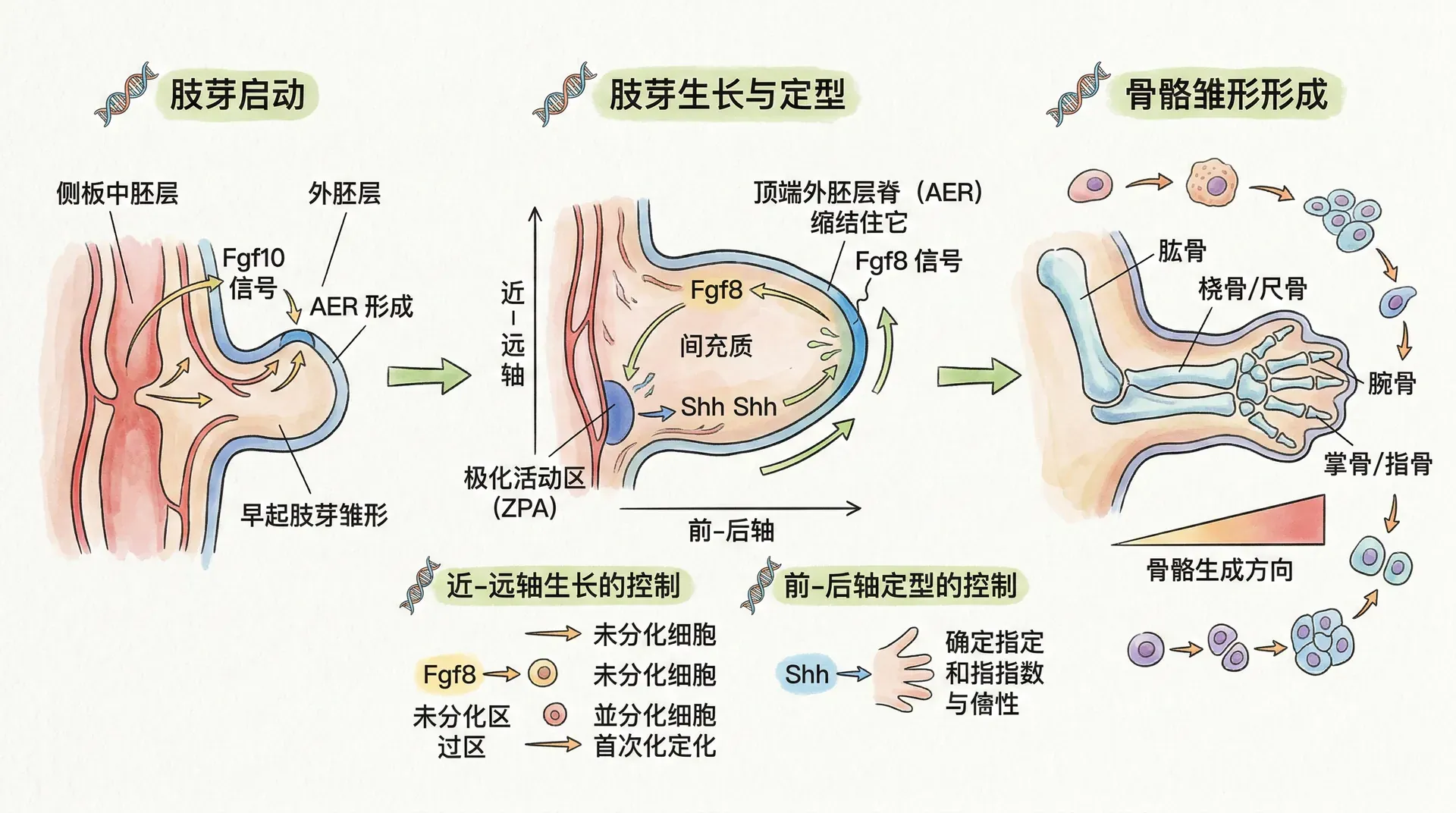
四肢的起始发生于侧板中胚层(lateral plate mesoderm)的局部膨出。大约在人类胚胎发育的第4周末,前肢肢芽出现在颈胸交界处,后肢出现在腰骶部,对应位置的侧板中胚层细胞快速增殖、外突,形成初期肢芽结构。肢芽的解剖结构包括内核的中胚层和外层包裹的外胚层。
在肢芽远端,外胚层显著增厚,形成顶外胚层嵴(AER)。AER像一条帽沿覆盖于肢芽的最前缘,是调控四肢生长的重要信号中心。AER主要分泌成纤维细胞生长因子(FGF),对下方中胚层产生持续的增殖及存活信号。如果实验性去除AER,则肢芽近远轴方向的增长会中断。
三个体轴的建立
四肢空间结构的分化,依赖于三个体轴的建立:
- 近远轴(proximodistal):从身体(肩/髋)到指趾;
- 前后轴(anteroposterior):从拇指/大脚趾到小指/小脚趾;
- 背腹轴(dorsoventral):手背/足背到手心/足底。
各轴的关键信号如下:
前后轴的模式形成主要依赖“极化活性区”(ZPA)。ZPA位于肢芽后缘,分泌Sonic Hedgehog(Shh)信号,形成后高前低的浓度梯度。实验表明,将ZPA组织移植到肢芽前缘,会形成镜像对称的双小指肢体,经典“ZPA移植实验”曾广泛用于发育机制研究。
背腹轴由外胚层发出的拮抗信号决定。背侧外胚层表达Wnt7a,诱导下方中胚层分化为背侧组织(如手背),间接激活Lmx1基因;腹侧外胚层表达En1,可抑制背侧信号,帮助确立手心身份。
以下总结四肢发育中三大信号中心的特征:
指趾的形成与雕刻
当肢芽发育至一定阶段后,其末端膨大并逐渐形成扁平的“手板”和“足板”,此时指趾之间尚无清晰间隙。指与趾的分离,是如下两种机制共同作用的结果:
- 局部增殖:未来指(趾)区域的细胞获得高生存及增殖信号,形成软骨基本模板。
- 指间细胞凋亡:手板和足板之间,指间区域主动触发程序性细胞死亡(apoptosis),逐步清除多余细胞,最终塑造出分明的五指(五趾)。
如下事例说明该过程的重要性:指间细胞凋亡缺失——在某些遗传背景或环境因素影响下,指间凋亡受阻,导致并指畸形(如部分江南地区“鸭蹼手”高发)。
骨骼成型机制举例
肢体的骨骼通常通过“软骨内成骨”完成。即:
- 中胚层细胞聚集为软骨细胞,形成长骨雏形。
- 血管侵入软骨,成骨细胞取代软骨并开始沉积骨基质。
- 骨生长板在骨骺端持续参与骨骼的线性生长。
器官发生中的细胞信号转导
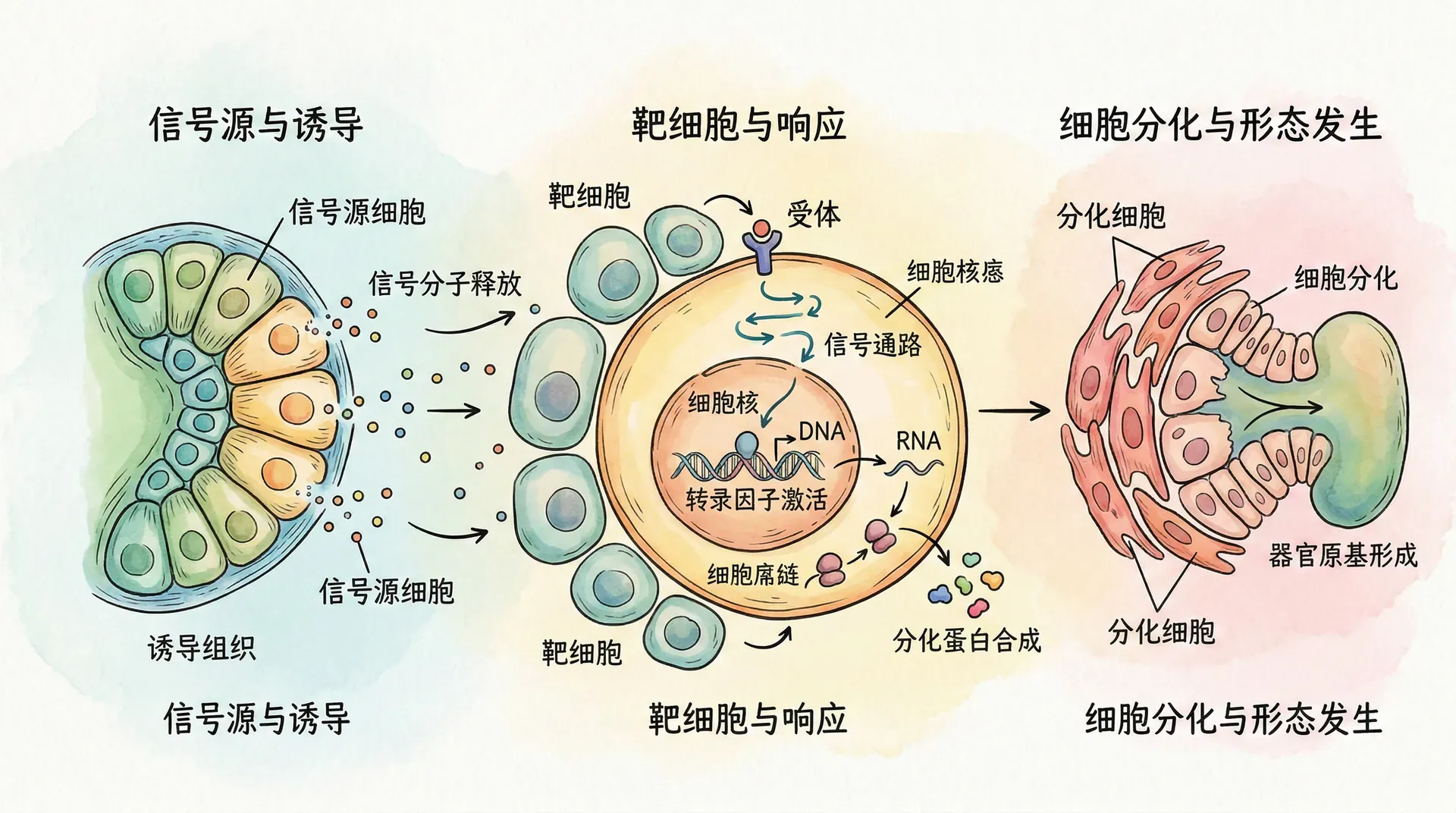
器官形成过程中,细胞间需要通过信号分子交流,实现精确的分工协作。这个过程称为“细胞信号转导”。发育生物学中,几条经典信号通路(Hedgehog、Wnt、FGF等)反复在不同时间、空间与器官类型中组合使用。
Hedgehog信号通路
- 该通路控制:神经管背腹极化、四肢前后轴分化、毛囊等器官形成。
- 基本机制为:Shh与受体Patched结合→解除Smoothened抑制→激活Gli转录因子→调控靶基因表达。
- 例子举要:神经管发育时,Shh形成从腹到背的浓度梯度。高浓度诱导运动神经元区分化,中低浓度形成中间和背侧神经元。
Wnt信号通路
Wnt信号广泛参与胚胎、干细胞和器官再生等诸多过程。以β-连环蛋白为核心,在无Wnt信号时降解β-catenin,有Wnt信号则激活其积累并调控相关基因。
FGF信号通路
FGF家族包含众多成员,可以激活MAPK、PI3K等多条下游信号通路,在胚胎发育过程中具有高度的时空调控特异性。例如,在四肢发育时,顶外胚层嵴(AER)分泌FGF8以维持中胚层的增殖;肺分支形成过程中,间充质释放FGF10以诱导上皮产生分支出芽;而在神经管发育中,FGF8参与不同脑区边界的划分,帮助神经系统结构化。
FGF信号的作用效果受到多种因素影响,包括信号分子的浓度、受体在细胞表面的表达水平,以及负反馈等调控机制。这些因素共同决定了细胞对FGF信号的响应强度与生物学效应。
信号通路的整合互作
多种信号通路间普遍存在复杂的互作及正负反馈。例如:
正反馈环路: 在肢芽发育中,ZPA分泌Shh促使AER维持FGF表达,反之FGF又维持ZPA活性,此互作强化局部信号,维持肢芽生长。
信号平衡: 肠道中Wnt、BMP和Notch分别调控干细胞增殖、分化和细胞命运平衡,构成维持肠上皮功能的“信号三角”。
下面信息展示Shh、Wnt、FGF信号在组织中的浓度梯度与响应分布:
上图体现三类关键信号在发育中的梯度分布模式。正是这些信号浓度与组合的差异,使细胞能够识别自己的空间位置,激活特定基因表达,从而行使相应分化命运。
发育缺陷与出生缺陷预防
胚胎发育虽然调控精密,但其高度复杂性使其易受遗传和环境因素干扰,从而诱发多种发育缺陷。了解这些成因和制定相应预防对提升人口健康水平意义重大。
主要器官缺陷与致畸因素
常见的出生缺陷包括神经管缺陷(如无脑畸形、脊柱裂)、先天性心脏病及肢体畸形等。神经管缺陷多见于中国北方,与母体叶酸缺乏密切相关;先天性心脏病在全国发病率较高,包括室间隔缺损、房间隔缺损等,部分可自愈,严重时需外科干预。肢体畸形则包括多指、并指及罕见的海豹肢症,后者曾因沙利度胺药物广泛使用引发关注,并促进了药物致畸性检测的发展。
发育缺陷的成因结合了遗传(如基因突变、染色体异常)与环境(药物、感染、营养、物理等)因素。不同发育时期对致畸因素的敏感性不同:受精后两周为“全或无期”,第三至第八周为器官形成关键期,对致畸因素最敏感,第八周后致畸影响主要体现在功能与生长方面。
三级预防策略及再生医学进展
中国采纳三级防控体系减少出生缺陷:一级预防着重孕前与孕早期的健康检查、叶酸补充、疫苗接种以及避免有害因素(如2009年起全国推行的免费优生健康检查);二级预防依赖产前筛查(超声、母血清学及无创基因检测)、高风险者通过羊穿、脐穿、绒毛膜活检等获得产前确诊并干预;三级预防则是新生儿期的疾病筛查、早期干预,典型项目有苯丙酮尿症、甲低及听力障碍筛查。
生物医学进步推动了发育缺陷干预向精准化迈进。CRISPR基因编辑已在动物模型中矫正在遗传疾病中的致病突变,体细胞和成体干细胞的基因治疗正进入临床阶段。干细胞与组织工程技术为再生和修复损伤器官带来新契机,如iPSC技术和3D生物打印心脏组织的创新成果。中国科研团队在iPSC建立、疾病建模及生物打印等领域均作出突出贡献。
上图呈现中国近年出生缺陷发生率的变化趋势:随着预防策略的广泛落实,尤其是叶酸普及,神经管缺陷显著下降;而先天性心脏病检出率受诊断技术发展先上升后随干预措施逐步降低。
总结
器官发生是一个高度精细的生物过程,涉及细胞的增殖、分化及精确定位。神经系统、心血管系统和四肢等器官在发育过程中体现了遗传调控与环境信号的复杂协同。每一个器官的形成都经历了多阶段的形态变化和功能塑造,确保了生物体结构和功能的完善。
在神经系统发育中,神经管的闭合与神经嵴细胞的迁移决定了中枢和周围神经结构的建立。心血管系统起源于原始心管,经过一系列的折叠和分隔,最终发育为结构复杂的心脏和循环体系。四肢的形成则依赖于信号分子的梯度分布和肢芽各区域的协调发育,精确塑造出手足的形态和前后、背腹等轴向结构。
近年来,分子机制的研究不断揭示器官形成过程中各种信号通路和基因网络的调控作用,如Notch、Wnt、Shh等信号在组织模式建成中的关键功能。干细胞及再生医学的发展也为受损组织和器官的修复开辟了新途径。随着对发育过程和病理机制理解的加深,针对先天性缺陷和组织损伤的预防与治疗策略得到了不断完善,为临床应用提供了理论基础和技术支持。
本节练习
1. 关于神经管形成的描述,下列哪项是正确的?
A. 神经管从头端开始闭合,向尾端延伸
B. 神经管闭合从中部开始,向头尾两端延伸
C. 神经管闭合从尾端开始,向头端延伸
D. 神经管各部分同时闭合
2. 在四肢发育中,极化活性区(ZPA)主要通过分泌哪种信号分子来控制前后轴的模式形成?
A. FGF8
B. Wnt7a
C. Sonic Hedgehog(Shh)
D. BMP4
3. 原始心管在发育过程中发生弯曲和扭转的主要原因是:
A. 心包腔的快速扩张
B. 心管的生长速度快于心包腔的扩张
C. 心肌细胞的不均匀收缩
D. 外部压力的作用
4. 下列关于神经嵴细胞的描述,哪项是错误的?
A. 神经嵴细胞来源于神经板和表皮外胚层的交界处
B. 神经嵴细胞具有广泛的迁移能力
C. 所有神经嵴细胞都分化为神经元
D. 黑色素细胞来源于神经嵴细胞
5. 胚胎发育中对致畸因素最敏感的时期是:
A. 受精后0-2周
B. 受精后3-8周
C. 受精后9-12周
D. 受精后13-16周
6. 请解释顶外胚层嵴(AER)在四肢发育中的作用,并说明如果AER功能缺失会导致什么后果。
7. 中国实施的出生缺陷三级预防策略是什么?请分别说明每一级预防的主要内容和意义。