行为的生物学基础
北京某个夜晚,一名30岁的女士独自走在昏暗的胡同里。突然,一名持刀歹徒上前威胁,但奇怪的是,她完全没有产生任何恐惧反应——心率正常,呼吸平稳,表情镇定。这种异常的冷静让歹徒不知所措,最终落荒而逃。
这名女士童年时期患过一种罕见疾病,损伤了大脑中控制恐惧的关键区域——杏仁核。杏仁核负责恐惧体验,它的缺失让她完全丧失了恐惧感。这个案例为科学家研究大脑与情感的关系提供了珍贵证据。这个案例生动展示了大脑对人类行为的精确控制。她能感受除恐惧以外的所有情感,证明了行为生物学基础的复杂性与功能专一性。
值得注意的是,无法感受恐惧并非好事。缺乏恐惧感意味着在判断危险时存在根本性的缺陷,当事人往往会在浑然不觉中将自己置于危险境地。
神经系统中的信息传递
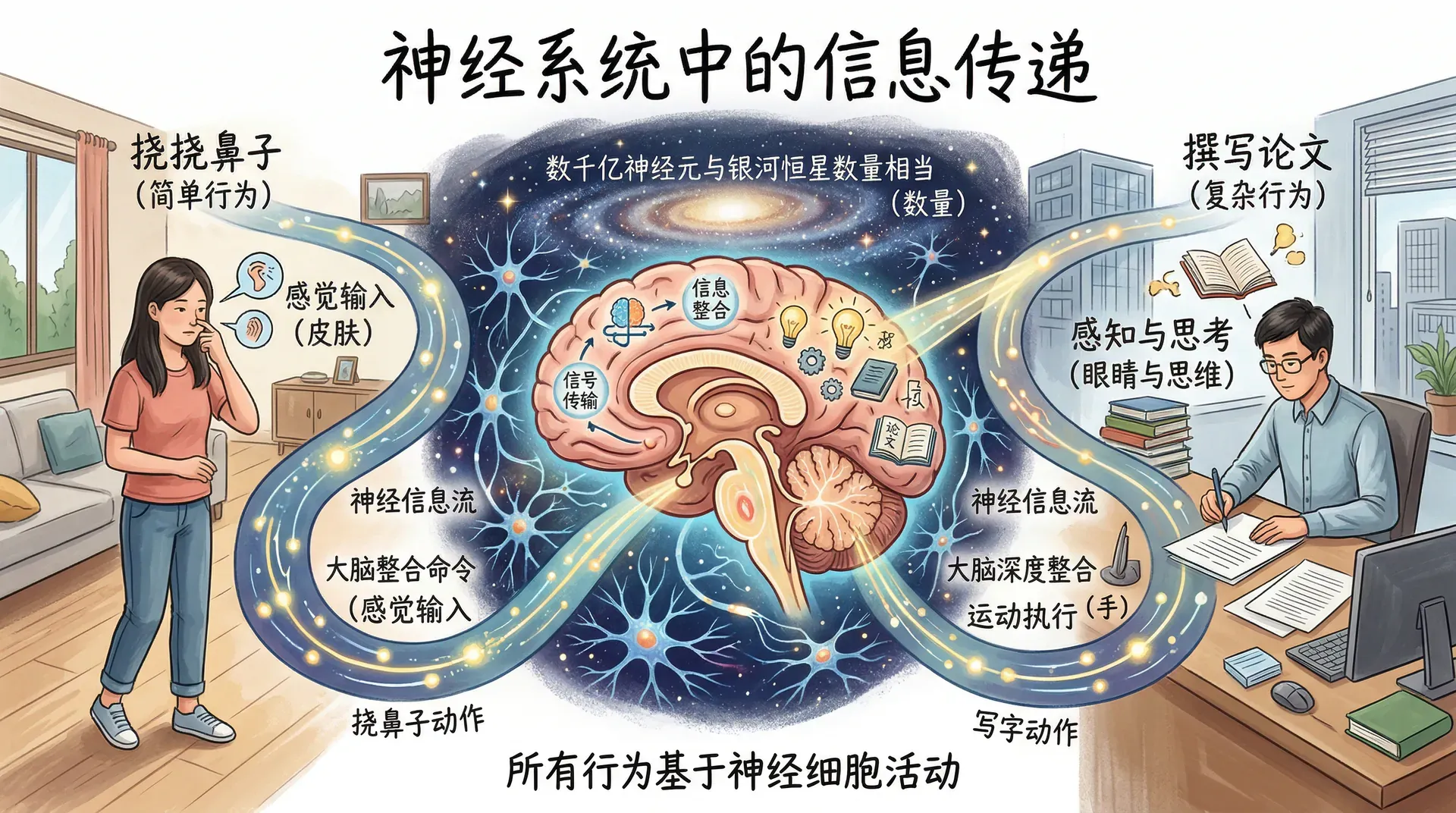
人类大脑至今没有任何计算机能够接近复制。神经系统包含的细胞数量与银河系中的恒星数量大致相当,这些细胞持续不断地整合和传递信息。无论是挠挠鼻子,还是撰写一篇论文,所有行为都以神经细胞的活动为基础。
当一名学生在备考期间观看悬疑剧时,紧张的情节让手心出汗、心跳加速。表面上只是在“看电视”,体内却正进行着极其复杂的神经活动:屏幕光线刺激眼睛,大脑瞬间解释视觉信息,神经信号快速传遍全身,触发一系列生理反应。这个日常场景揭示了行为对快速信息处理的依赖——信息从眼睛传到大脑,从大脑传到肌肉,又从手掌传回大脑。神经系统本质上是一个复杂的通信网络,信号在其中不断地被传输、接收和整合。
神经元
神经系统由细胞组成,其中的细胞分为两大类:神经元和胶质细胞。神经元是神经系统中接收、整合和传输信息的单个细胞,是允许神经系统内部通信的基本连接单位。绝大多数神经元只与其他神经元通信,少数神经元从感觉器官接收外部信号,或将信息从神经系统传递到肌肉。
一个典型神经元包含以下基本结构。细胞体包含细胞核和大多数细胞共有的化学机器。神经元的其余部分专门用于处理信息:神经元有许多分支状的树突,专门负责接收来自其他细胞的信号;树突将信息汇入细胞体,再沿轴突向外传导。轴突是一根长而细的纤维,将信号从细胞体传输到其他神经元或肌肉与腺体,有时长达数十厘米甚至更长。
在人类中,许多轴突被包裹在含有高浓度白色脂肪物质的鞘膜中,称为髓鞘。髓鞘能够加速沿轴突传播的信号速度。多发性硬化症患者丧失肌肉控制,正是由于髓鞘退化导致信号无法有效传输所致。轴突末端是一簇末端按钮,这些小结节分泌称为神经递质的化学物质,充当信使激活附近的神经元。神经元相互连接的点称为突触,是信息从一个神经元传递到另一个神经元的关键连接点。
信息在树突处被接收,经过细胞体和轴突传递,最终在突触这一连接点传输到其他细胞的树突。这条信息通路是所有神经系统活动的根本基础。
胶质细胞
胶质细胞遍布整个神经系统,为神经元提供各种类型的支持。胶质细胞通常比神经元小得多,但在数量上超过神经元约十比一,占大脑体积的一半以上。胶质细胞承担着多项重要功能:为神经元提供营养,帮助清除神经元的代谢废物,并在许多轴突周围提供绝缘保护。包裹某些轴突的髓鞘正是由特殊类型的胶质细胞产生的。在人类胚胎神经系统的发育过程中,胶质细胞同样扮演着复杂而不可或缺的角色。
近年来的研究表明,胶质细胞可能也会发送和接收化学信号。某些类型的胶质细胞能够检测神经冲动,并向其他胶质细胞发出信号。神经科学家目前正努力厘清这套信号系统如何与神经通信系统协同运作。一些早期发现颇为引人入胜:研究显示胶质细胞可能在记忆形成中发挥重要作用,而胶质组织的逐渐退化则可能是阿尔茨海默病的诱因之一。
神经冲动
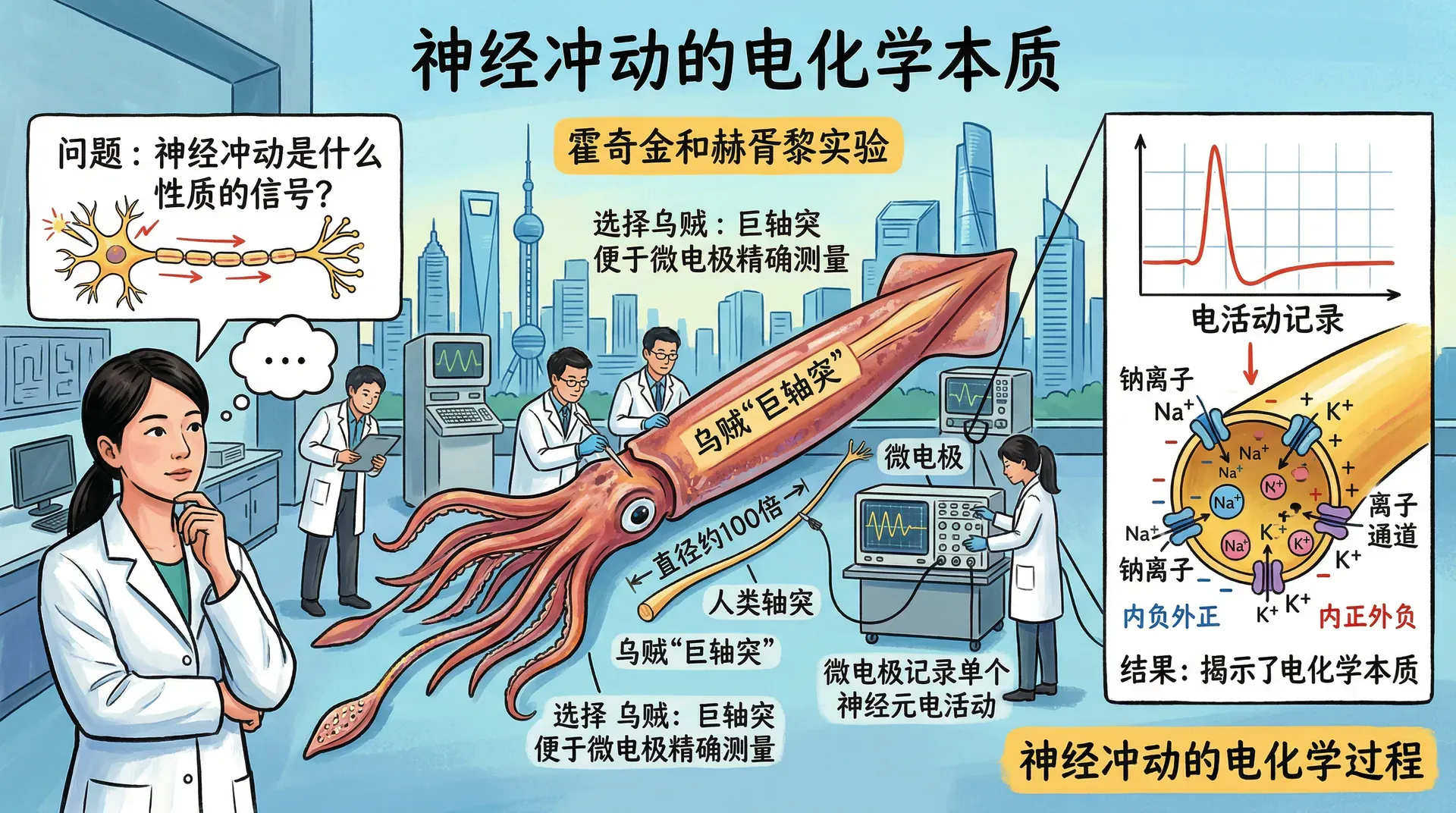
当神经元受到刺激时究竟发生了什么?沿轴突传播的神经冲动,本质上是什么性质的信号?这些问题由霍奇金和赫胥黎在针对乌贼轴突的开创性实验中首先得到解答。他们选择乌贼,是因为乌贼拥有一对直径约为人类轴突百倍的“巨轴突”,更便于放置微电极进行精确测量。通过记录单个神经元的电活动,他们揭示了神经冲动的电化学本质。
静息状态的神经元
神经元内外都充满含有带电原子和分子的液体,这些带电粒子称为离子。带正电的钠离子和钾离子,以及带负电的氯离子,以不同速率穿越细胞膜流动。流速的差异导致细胞内负电荷离子的浓度稍高于细胞外,由此产生的电压使静息状态的神经元犹如一枚小电池,储存着潜在的能量。
静息电位是细胞不活跃时的稳定负电荷,约为负70毫伏,大约相当于一节手电筒电池电压的二十分之一。这个电位是神经元随时准备响应刺激的基础状态。
动作电位
只要神经元的电压保持恒定,细胞就处于静息状态,不发送任何信息。当神经元受到刺激时,其细胞膜中的通道打开,短暂允许带正电的钠离子涌入。刹那间,神经元的电荷从负变正,产生所谓的动作电位。动作电位是神经元电荷的极短暂变化,并沿轴突向下传播——就像一根导火索被点燃后,火花沿着导火索依次蔓延,电压变化也以同样的方式沿轴突逐段推进。
在动作电位触发后,允许钠离子进入的细胞膜通道随即关闭,需要短暂时间才能重新准备好。在此期间,神经元无法触发新的动作电位,这段时间称为绝对不应期,通常只有1至2毫秒。随后是短暂的相对不应期,神经元可以再次被触发,但触发阈值升高,需要更强的刺激才能启动新的动作电位。
全或无定律
神经冲动遵循“全或无”定律,类似于扣动扳机的动作——要么完全发射,要么完全不发射,没有中间状态。较弱的刺激不会产生较小的动作电位,神经元触发的动作电位大小始终相同。尽管如此,神经元仍然能够传达刺激强度的信息,方式是改变触发动作电位的频率——较强的刺激会使细胞以更快的速率产生连续神经冲动。
各种神经元以不同的速度传输冲动,较粗的轴突传输速度更快,最高可达每秒100米,相当于每小时360公里,整个复杂的神经传输过程仅需几千分之一秒。
突触
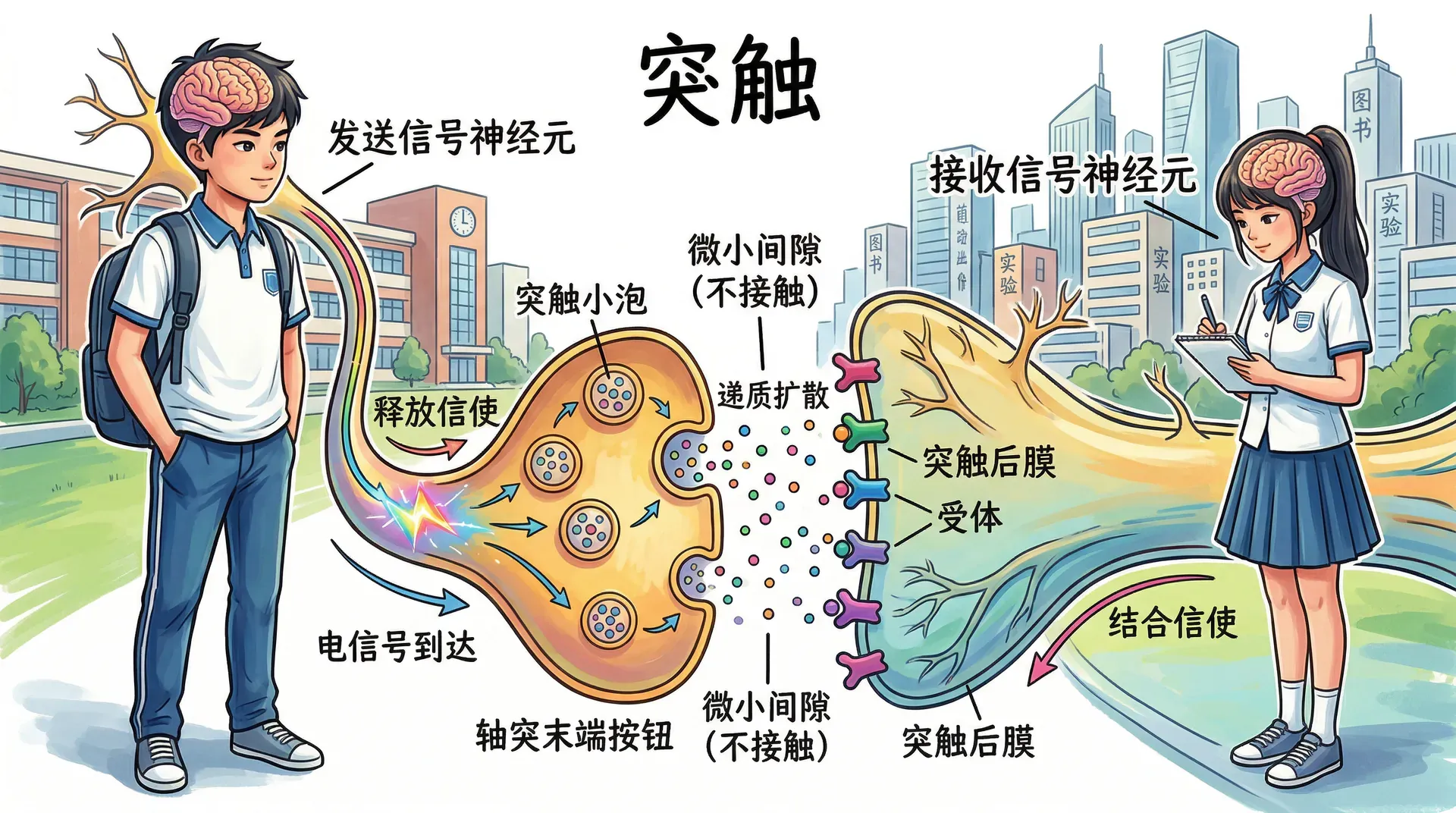
在神经系统中,神经冲动作为信号在神经元之间传递,而这种传递发生在称为突触的特殊连接处,依赖化学信使来完成跨越细胞间隙的信号传导。
突触的结构中,两个神经元实际上并不直接接触,而是被一个极微小的突触间隙分开。发送信号的神经元称为突触前神经元,接收信号的神经元称为突触后神经元。当动作电位到达轴突末端按钮时,触发神经递质的释放。这些化学物质储存在称为突触小泡的小囊中,小泡与突触前细胞膜融合,将内容物溢入突触间隙,神经递质随后扩散至接收细胞的膜面,并与突触后细胞膜上的受体分子结合。
当神经递质与受体结合时,细胞膜上的反应会产生突触后电位,即突触后细胞膜上受体位点的电压变化。突触后电位不遵循全或无定律,其大小会有所不同,且按比例增加或减少接收细胞产生神经冲动的概率。信息在细胞间的传递分为两种性质:兴奋性突触后电位是正电压变化,增加突触后神经元触发动作电位的可能性;抑制性突触后电位是负电压变化,降低突触后神经元触发动作电位的可能性。
兴奋性或抑制性效应只持续几分之一秒,随后神经递质从受体位点漂离,或被酶代谢为非活性形式。大多数神经递质通过重摄取被重新吸收到突触前神经元中,这一过程使突触能够回收其化学材料,为下一次信号传递做好准备。
神经网络
一个神经元可能同时从数千个其他神经元接收信号,同样也可能将自身的信息传递给数千个神经元。因此,神经元必须完成的工作远不止简单地转发信息——它必须整合来自众多突触的输入信号,然后“决定”是否触发神经冲动。
如果细胞内积累了足够多的兴奋性突触后电位,电流叠加使电压达到阈值,动作电位便被触发。然而,若同时也有许多抑制性突触后电位发生,它们将抵消兴奋性信号的效果。神经元的状态,始终是兴奋性与抑制性影响之间动态的加权平衡。数百万神经元必须协调触发,才能产生最微不足道的一个念头。大多数神经元在复杂的链条、通路、回路和网络中相互连接,我们的感知、思想和行为依赖于精细神经网络中神经活动的整体模式。这些网络中的连接是流动可变的——新突触连接可能形成,旧连接也可能萎缩。突触的消除似乎比新突触的创建在神经网络的塑造中发挥更大的作用:神经系统通常先形成比所需更多的突触,再逐渐修剪掉不太活跃的连接。
神经递质与行为
神经系统依赖化学信使在神经元之间传递信息,这些神经递质是行为的生化基础,在从肌肉运动到情绪调节、心理健康的方方面面都发挥着关键作用。目前已确定的经典小分子神经递质约有九种,此外还有约40种神经肽化学物质至少在部分情况下充当神经递质,以及各种新近认识的信号分子。
特定的神经递质在特定类型的突触处工作。神经递质与受体位点的结合过程很像锁和钥匙的关系——只有当神经递质的形状与受体位点匹配时,结合才能发生。这种高度特异性减少了密集神经元之间的信号串扰,使神经系统的通信更加精确可靠。
乙酰胆碱
细胞通过释放化学物质进行通信的发现,最初正是在乙酰胆碱这种神经递质上取得的。乙酰胆碱在整个神经系统中广泛存在,是运动神经元与随意肌肉之间唯一使用的神经递质。人体的每一个动作——打字、行走、说话、呼吸——都依赖于运动神经元向肌肉释放的乙酰胆碱。
乙酰胆碱似乎也参与注意力、觉醒和记忆的维持。大脑某些区域乙酰胆碱供应不足与阿尔茨海默病的记忆丧失有关。目前用于延缓阿尔茨海默病进展的药物,其主要机制正是通过增强乙酰胆碱的活性来实现,尽管效果相对有限。
乙酰胆碱的活性可能受到其他化学物质的影响。突触受体位点有时会被外来化学物质“欺骗”。尼古丁就是一个典型案例:它与乙酰胆碱的受体位点结合,产生与乙酰胆碱相似的突触后电位。在专业术语中,尼古丁是乙酰胆碱的激动剂——即模仿神经递质作用的化学物质。与之相对,拮抗剂是占据受体位点却不产生突触后电位的化学物质,通过阻断受体位点来对抗神经递质的正常作用。
当前流行的肉毒杆菌美容注射,其作用原理是阻断注射部位附近运动神经元与肌肉之间的乙酰胆碱受体,从而暂时麻痹肌肉、防止皱纹形成。但这种效果只能持续约三至五个月,因为突触会逐渐产生新的乙酰胆碱受体来适应变化。
单胺类神经递质
单胺类神经递质包括多巴胺、去甲肾上腺素和血清素三种,使用这些神经递质的神经元调节着日常行为的许多方面。
多巴胺被控制随意运动的神经元大量使用。大脑特定区域的多巴胺神经元退化会导致帕金森病,这种疾病以震颤、肌肉僵硬和随意运动控制减弱为主要特征。用于治疗帕金森病的左旋多巴等药物,在大脑中可转化为多巴胺,以部分补偿多巴胺活性的不足。
血清素系统的神经元与睡眠和觉醒的调节以及饮食行为密切相关。大量研究表明,使用血清素的神经回路参与调节攻击行为,血清素回路的异常也被认为是饮食障碍和强迫症的诱发因素之一。大脑中单胺类神经递质的异常水平与多种心理障碍的发生有关,患有抑郁症的人在去甲肾上腺素和血清素突触处的激活水平普遍降低。多巴胺假说则认为,多巴胺突触的异常活动在精神分裂症的发展中起着关键作用——这种严重精神疾病影响约1%的人口,以非理性思维、幻觉和现实接触障碍为特征,研究表明多巴胺回路的过度活跃可能是其神经化学基础的重要组成部分。
安非他明和可卡因等兴奋性毒品的强烈效应,也主要通过在多巴胺和去甲肾上腺素突触处制造活动风暴来实现。这一发现支持了部分理论家的观点:大多数成瘾性物质的奖励效应,都依赖于特定多巴胺通路中活动的异常增加,而这条多巴胺通路的功能失调,也被认为是药物渴求和成瘾的核心机制之一。
氨基酸神经递质
另一类重要的神经递质由氨基酸组成。伽马氨基丁酸几乎在所有存在的突触处都发挥抑制作用,是大脑中抑制功能的主要执行者。其他神经递质如乙酰胆碱和去甲肾上腺素,可以根据结合的突触受体类型产生兴奋性或抑制性效应,而伽马氨基丁酸的效应几乎一成不变地指向抑制。
伽马氨基丁酸受体广泛分布在大脑中,估计存在于约40%的突触中。研究表明,伽马氨基丁酸参与人类焦虑情绪的调节,其回路的功能紊乱可能导致某些类型的焦虑障碍。谷氨酸是另一种广泛分布的氨基酸神经递质,与伽马氨基丁酸的纯粹抑制性不同,谷氨酸始终发挥兴奋作用,其最重要的功能在于对学习和记忆的贡献。近年来,谷氨酸回路的功能紊乱被认为可能与精神分裂症的某些症状有关。
内啡肽
20世纪70年代,中国科学家韩济生在研究针刺镇痛的神经化学机制时,为揭示大脑内源性镇痛物质做出了重要贡献。当时,科学界已初步发现吗啡通过与大脑中的特定受体结合发挥作用,这一发现随即引发了一个核心问题:为什么大脑会配备吗啡受体?研究人员意识到,神经系统必然存在自身产生的类吗啡物质。这些物质后来被命名为内啡肽,意为“内部产生的类阿片化学物质”。
随后的研究证实,内啡肽及其受体广泛分布于人体各处,主要参与疼痛的调节,同时也影响饮食行为和机体对压力的反应。内啡肽的发现催生了关于疼痛与快乐神经化学基础的新理论。研究人员推测,人体的天然内啡肽也可能具有产生愉悦感的能力,这或许可以解释为什么长跑运动员有时会体验到“跑步者的愉悦感”——长跑引起的身体疼痛可能触发内啡肽的释放,这些内啡肽在中和部分痛感的同时,带来一种轻快的兴奋感。
神经系统的组织
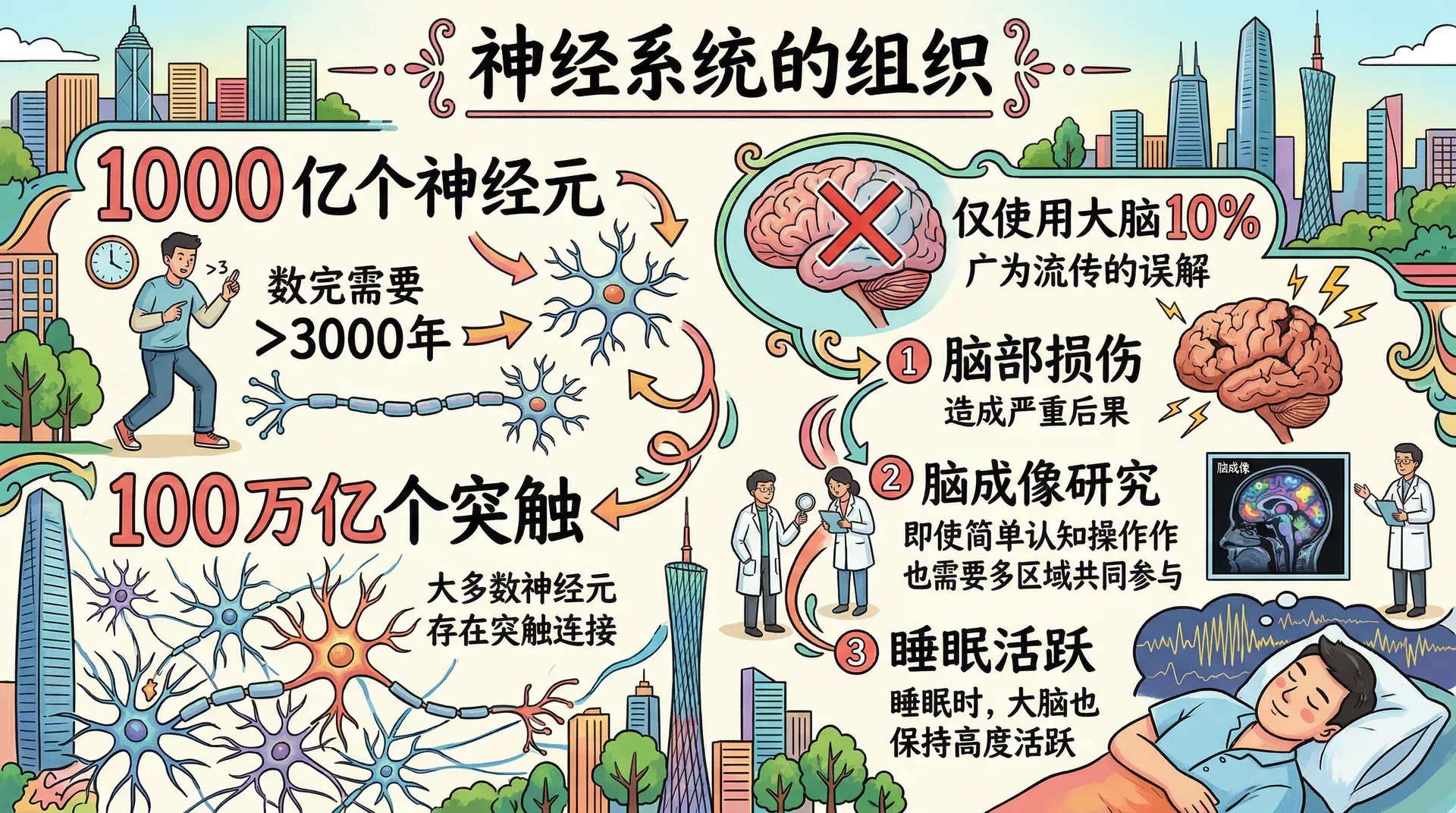
人类大脑中大约有1000亿个神经元,如果以每秒数一个的速度不停计数,需要超过3000年才能数完。而且大多数神经元与许多其他神经元存在突触连接,人脑中的突触总数可能高达100万亿个。如此庞大的数字,催生了“人们只使用了大脑10%”这一广泛流传的说法,但这完全是误解。如果大脑真有90%是冗余的,那么脑部损伤就不会造成如此严重的后果;脑成像研究清楚地表明,即便是简单的认知操作,也需要大脑多个区域共同参与。实际上,即使在睡眠期间,大脑也保持着高度活跃的状态。
神经系统从整体上可分为中枢神经系统和周围神经系统两大部分,二者协同工作,共同维系着人体所有的生理和心理活动。
周围神经系统
周围神经系统由位于大脑和脊髓以外的所有神经组成,是中枢神经系统与身体其余部分之间的通信通道。周围神经系统进一步分为躯体神经系统和自主神经系统两个主要子系统。
躯体神经系统
躯体神经系统由连接到随意骨骼肌和感觉受体的神经组成,让人得以感受外部世界并在其中自由活动。这些神经承担着双向传递的任务:传入神经纤维将皮肤、肌肉和关节的感觉信息向内传递到中枢神经系统,传出神经纤维则将中枢神经系统的运动命令向外传达到身体各处的肌肉。每条躯体神经都同时包含这两类纤维,犹如一条兼具上行与下行车道的双向公路。
自主神经系统
自主神经系统控制着人们通常不会有意识地去调节的内脏功能,如心率、消化和出汗,由连接到心脏、血管、平滑肌和腺体的神经组成。虽然“自主”意味着这个系统能够相对独立地运作,但它最终仍受中枢神经系统的调控。
试想一个上班族在深夜独自回家,发现家门半开、门锁被撬坏。怀疑家中遭到入室盗窃,他的心率和呼吸会立刻加快,血压迅速升高,身上起鸡皮疙瘩,手掌开始出汗。这些难以主观控制的生理反应,正是自主神经系统激活的典型表现。心理学家沃尔特·坎农将这种反应命名为“战斗或逃跑反应”。
自主神经系统的交感分部负责动员身体资源应对紧急情况,产生战斗或逃跑反应——它会抑制消化过程,将血液从外围输送到核心肌群,并向肾上腺发送信号,触发激素释放,使身体为行动做好准备。与之相对,副交感分部负责保存身体资源,激活允许身体节约和储存能量的过程,如减慢心率、降低血压并促进消化。两个分部相互制衡,共同维持身体在不同状态下的动态平衡。
中枢神经系统
中枢神经系统由大脑和脊髓组成,受到头骨的坚硬保护,并被称为脑膜的包围鞘进一步守护。此外,中枢神经系统还沐浴在脑脊液中,这种特殊液体滋养着大脑,同时为其提供缓冲保护。大脑中充满脑脊液的空腔称为脑室。
脊髓
脊髓是中枢神经系统的重要组成部分,从大脑底部延伸至腰部以下,通过周围神经系统将大脑与身体的其余部分相连接。它承载着大脑向外传达命令的运动轴突,以及将来自全身感觉信息传入大脑的感觉轴突。临床上许多形式的瘫痪都由脊髓损伤引起,这一事实深刻揭示了脊髓在整个神经通信体系中不可替代的地位。
大脑
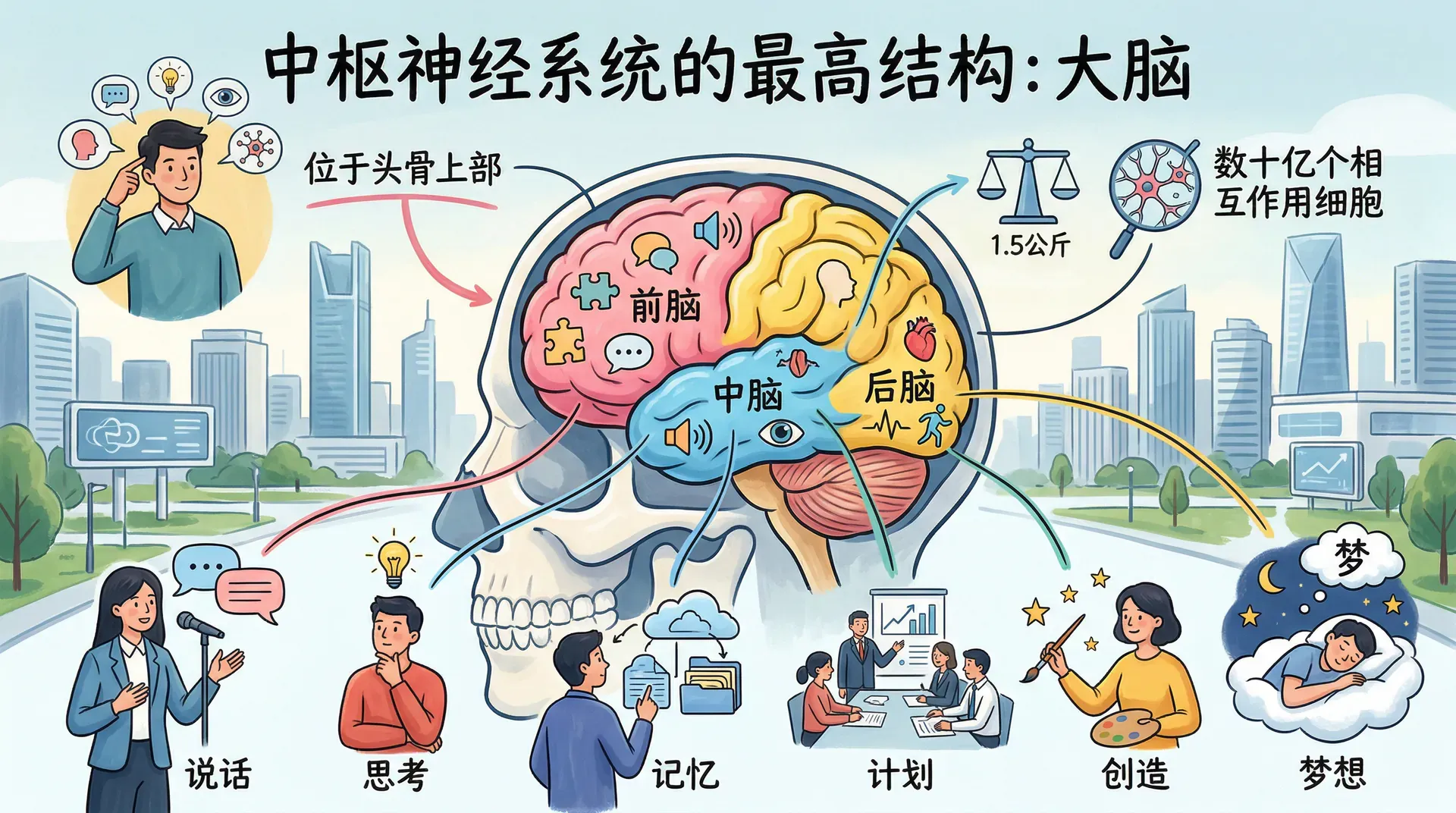
中枢神经系统的最高结构是大脑。从解剖学上讲,大脑是填充头骨上部的中枢神经系统部分。它重约1.5公斤,看似不大,却包含数十亿个相互作用的细胞。这些细胞整合来自身体内外的信息,协调身体的行动,使人类能够说话、思考、记忆、计划、创造和梦想。大脑可以分为三个主要区域:后脑、中脑和前脑。
后脑
后脑包括小脑和脑干下部的两个结构:延髓和脑桥。延髓连接脊髓,控制主要的无意识但极为重要的生命功能,包括血液循环、呼吸、肌肉张力的维持,以及调节打喷嚏、咳嗽和流涎等反射动作。脑桥包含连接脑干与小脑的纤维桥,并含有参与睡眠和觉醒调节的若干细胞体群。
小脑位于脑干后表面附近,是一个相对较大且深度折叠的结构,对运动协调和平衡感至关重要。肌肉运动的实际命令来自更高的大脑中心,但小脑在组织和整合指导这些运动的感觉信息方面扮演着核心角色。当你向一侧伸出手臂,再流畅地将指尖停在鼻尖上时,背后正是小脑的精确调控。饮酒后之所以难以完成这个简单动作,正是因为小脑是最早受到酒精抑制的脑结构之一。小脑损伤会破坏书写、打字或演奏乐器等精细运动技能。近年的研究还发现,小脑与前额皮质之间存在神经回路,这条回路参与注意力、计划和视觉感知等更高级的功能,表明小脑的作用远不局限于运动控制。
中脑
中脑是脑干中位于后脑和前脑之间的部分,包含一个整合视觉和听觉等感觉过程的关键区域。从中脑发出的重要多巴胺释放神经元系统,向大脑各处更高级的中枢投射,参与随意运动的执行。导致帕金森病的多巴胺合成下降,正是由于位于中脑的特定结构退化所致。
贯穿后脑和中脑的是网状结构,位于脑干的中央核心区域,参与调节肌肉反射、呼吸和疼痛感知。网状结构最广为人知的功能是调控睡眠与觉醒——其上行纤维的活动对维持清醒状态至关重要。
网状结构可以被理解为大脑的“总开关”,决定着意识的开启与关闭。当这一结构受到严重损伤时,人可能陷入持续的昏迷状态。
前脑
前脑是大脑中最大、最复杂的区域,包含丘脑、下丘脑、边缘系统和大脑等多种重要结构。丘脑、下丘脑和边缘系统形成前脑的核心,位于脑干顶部附近。大脑位于其上方,是复杂思维的中心,其褶皱的外表面即为大脑皮质。
丘脑
丘脑是前脑中的核心中继结构,所有感觉信息(嗅觉除外)都必须先经过丘脑,才能到达大脑皮质。丘脑由多个细胞体集群组成,每个集群负责将特定类型的感觉信息中继到皮质的相应区域。然而,将丘脑仅仅定位为被动的中继站并不准确,研究表明丘脑在整合来自不同感觉通道的信息方面同样发挥着积极的主动作用。
下丘脑
下丘脑是位于前脑底部的小型结构,尽管体积不比蚕豆大,却包含众多功能各异的细胞群,参与基本生物需求的调节。下丘脑的一项重要功能是控制自主神经系统,此外它还充当大脑与内分泌系统之间的关键接口。
下丘脑在调节与生存相关的基本生物驱力中发挥重要作用。动物实验中,切除下丘脑外侧区域的动物会完全丧失进食动机,即便食物近在眼前也会因不思饮食而饿死;而电刺激同一区域则会引发持续性的过度进食行为。这些结果清楚表明,下丘脑深度参与饥饿、口渴、性动机和体温调节等基本生理过程的调控。
边缘系统
边缘系统是一个松散连接的结构网络,大致位于大脑皮质和皮质下区域之间的边界地带,因此得名“边缘”。边缘系统并非一个边界分明的解剖结构,科学家对其确切范围至今存在争议,但广义上包括下丘脑、海马体、杏仁核、嗅球和扣带回等结构,共同参与情感、记忆和动机的调节。
海马体和相邻结构在记忆过程中扮演重要角色。一些理论认为,海马体负责将新信息转化为持久记忆代码的“巩固”过程。有充分证据将边缘系统与情感体验联系起来,其中杏仁核被认为在恐惧反应的学习以及基本情感反应的处理中处于核心位置。开篇描述的那位女士,正是因为杏仁核受损,才失去了感知恐惧的能力。
边缘系统也是大脑中“奖励回路”最为集中的区域之一。早期动物实验中,研究人员发现大鼠会不惜体力反复按压杠杆,只为向自身大脑特定区域施加电刺激,甚至在饥饿状态下也优先选择自我刺激而非进食,由此推断这些脑区与强烈的快感体验相关。内侧前脑束穿过下丘脑的区域似乎是最集中的自我刺激位点,该区域富含多巴胺释放神经元,成瘾性药物的愉悦效应在相当程度上也通过这条多巴胺回路的激活来实现。
大脑皮质与高级功能
大脑是人脑中最大、最复杂的部分,负责最高级的心理活动,包括学习、记忆、思考和意识本身。大脑皮质是大脑复杂折叠的外层,其表面积约为1500平方厘米,通过大量皱褶被容纳在有限的头骨空间内。大脑分为左右两个半球,被一条纵向裂隙分开,两个半球之间通过胼胝体——一条厚实的神经纤维束——相互连接和通信。每个半球进一步分为四个叶片,各有其功能侧重。
额叶是人脑中最大的叶片,控制肌肉运动的初级运动皮质就位于此。值得注意的是,运动皮质分配给不同身体部位的区域大小,并不取决于该部位的体积,而是取决于该部位运动的精细程度。手指、嘴唇和舌头获得了远超其体积比例的皮质面积,而大腿和肩膀这类进行粗糙运动的较大部位,反而只占据很小的皮质区域。
在额叶运动皮质前方区域,研究人员发现了“镜像神经元”——这类神经元在执行某个动作时被激活,在观察他人执行同样动作时同样被激活。镜像神经元系统被认为在模仿学习、理解他人意图以及产生移情能力等社会认知功能中发挥基础性作用。
前额皮质占据大脑皮质面积约三分之一,是多种高级认知功能的核心区域,包括工作记忆、关系推理以及某些类型的决策。其对工作记忆和关系推理的重要贡献,使部分理论家认为前额皮质可能包含某种“执行控制系统”,负责监控、组织和整合思维过程。
大脑的可塑性
长期以来,人们认为大脑解剖结构的重大变化只发生在发育早期,成年后便基本固定。然而,研究逐渐揭示,大脑的解剖结构和功能组织比人们原先设想的更具可塑性,能够随着经验而变化和重组。
研究发现,习练弦乐的音乐人,其体感皮质中接收左手手指输入的区域明显大于普通人,因为弦乐演奏需要持续用左手按弦。国内有研究对参加杂技团培训的学员进行追踪,发现连续数月训练后,学员在处理视觉和运动任务的脑区中出现了可观察到的结构变化。这些发现表明,特定的行为经验可以实实在在地改变大脑的结构。
当感觉通路受损或脑组织被破坏时,神经系统同样表现出显著的重组能力。在先天失明或因病丧失视力的个体中,通常专门用于视觉处理的枕叶区域被“征召”来协助进行语言和触觉信息处理。脑损伤后,健康神经元也会尝试代偿邻近受损神经元的功能。此外,成年大脑也可以产生新的神经元,这一发现颠覆了神经发生只发生在发育阶段的传统观念。研究已令人信服地证明,成人的嗅球和海马体能够持续形成新神经元。考虑到海马体在记忆和学习中的重要地位,神经发生可能在维持认知功能方面发挥着持续的作用。
经验可以塑造大脑结构的特征。弦乐演奏家的体感皮质、经过专项训练的杂技学员大脑,都在特定区域表现出可测量的结构差异,证明行为经验能够改变大脑的物理形态。
感觉通路的损伤或脑组织的破坏可以引发神经重组。失去某一感觉输入后,相应脑区会被其他感觉功能“征用”,体现了大脑在受损情况下的自我适应倾向。
成年大脑可以产生新的神经元。神经发生并不局限于发育早期,成人的海马体和嗅球中持续有新神经元生成,这一发现深刻改变了人们对大脑可塑性的理解。
大脑的可塑性并非无限,随年龄增长会逐渐减弱,且严重损伤后的恢复能力也相当有限。尽管如此,大脑神经回路所表现出的可塑性,是人类终身适应环境变化的重要生物学基础。
探索大脑内部
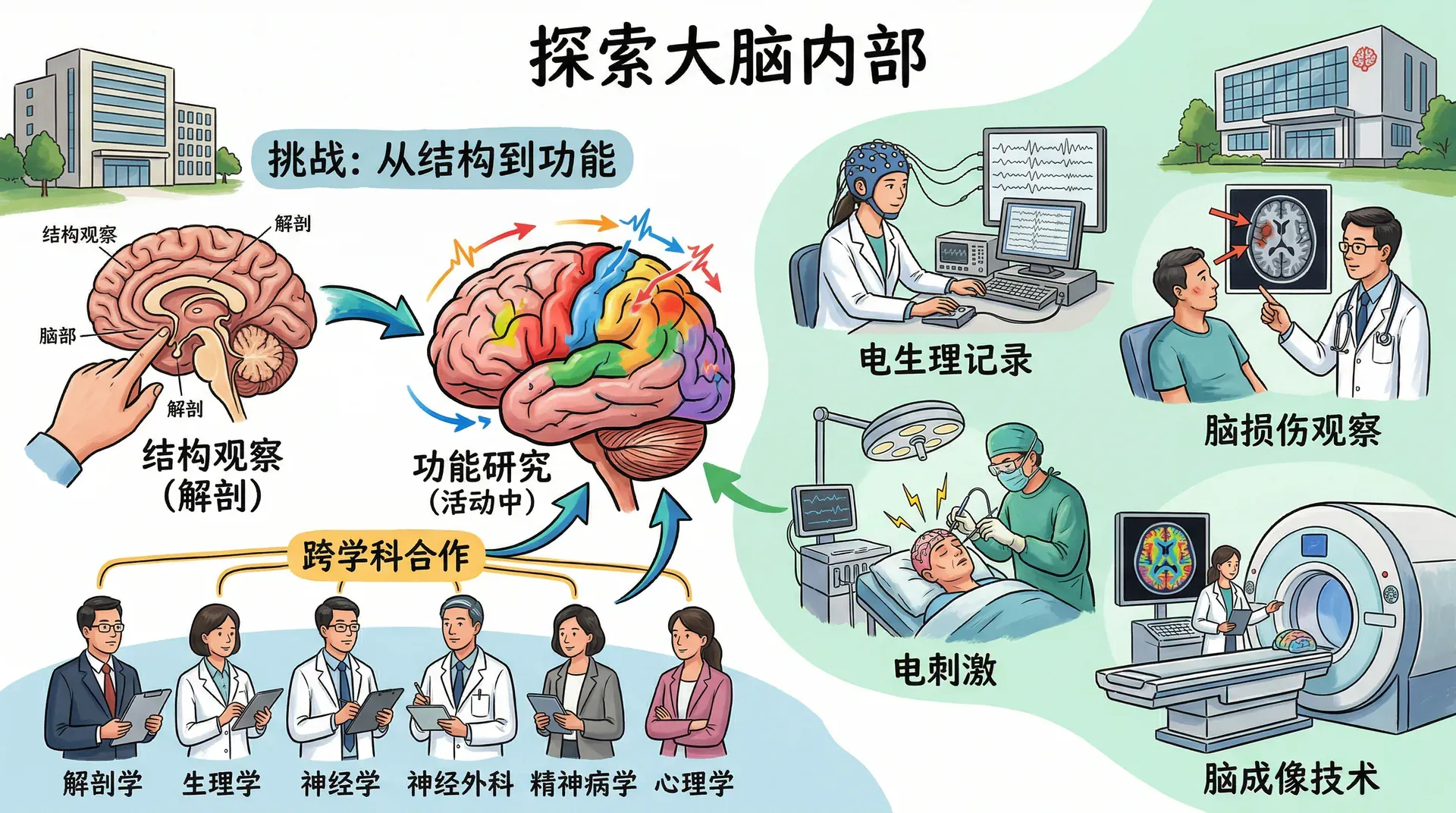
研究大脑各部分与行为之间关系的科学家面临着特殊的挑战——大脑的解剖结构可以通过解剖标本来观察,但要揭示大脑功能,则必须研究活动中的大脑。为此,神经科学家发展出了一系列专门的研究方法,涵盖电生理记录、脑损伤观察、电刺激和多种脑成像技术。
大脑研究通常涉及来自解剖学、生理学、神经学、神经外科、精神病学和心理学等多个学科的神经科学家之间的密切合作,这种跨学科的研究模式极大地推动了人类对大脑与行为关系的认识。
记录大脑的电活动
大脑的电活动可以通过脑电图来记录。这项技术由德国精神科医生汉斯·伯格于1929年发明,通过附着在头皮表面的记录电极,随时间监测大脑大范围的电活动模式。
通常将六到十个电极固定在头骨的不同位置,电极汇总并放大来自大脑内数千个脑细胞的电位,产生的记录被转换成称为“脑波”的线条图形。不同的脑波模式与不同的心理状态相对应,为研究者提供了大脑活动的整体概览。脑电图在临床上常用于脑损伤、癫痫和其他神经疾患的诊断。在研究领域,它被用于记录参与者从事特定活动或体验特定情绪时的大脑活动模式。在国内的睡眠研究领域,脑电图已成为探索睡眠神经基础不可或缺的工具。
观察受损大脑的影响
关于大脑与行为关系的许多重要见解,来自对特定脑区受损患者的行为变化的系统观察。这种研究路径的局限在于,可供观察的案例数量有限,受损位置和严重程度往往难以精确确定,且患者个人经历的差异会引入难以控制的额外变量。
为了更精确地研究特定脑区的功能,科学家有时在动物实验中通过人为手段损毁特定脑结构,再观察行为上的变化。损毁通常通过向目标脑区插入电极并通以高频电流来完成,配合立体定位仪实现对深部脑区的精确定位。这种动物实验方法在揭示大脑功能方面积累了大量宝贵的基础研究成果。
激活大脑区域
大脑电刺激是通过向大脑结构发送微弱电流来激活目标区域的技术。这种方法并非完全复制大脑中的自然信号,但通常足以激活植入电极的脑区,从而研究该区域的功能。大多数电刺激研究在动物身上进行,在人类身上的应用主要限于脑外科手术期间,用于术前“绘制”功能区地图,以避免手术损伤关键脑区。
经颅磁刺激
经颅磁刺激是一种新兴技术,允许科学家暂时性地增强或抑制大脑特定区域的活动。操作时,安装在小桨上的磁线圈被放置在受试者头部的目标区域,所产生的磁场能渗透到约两厘米的深度。通过调整磁脉冲的时间和持续时间,研究人员可以升高或降低目标脑组织中神经元的兴奋性。
这项技术的最大优势在于能以无痛、非侵入性的方式在人体上短暂制造“虚拟损伤”,既避开了伦理问题,又绕过了自然脑损伤研究中众多难以控制的变量。目前已被用于探索特定脑区在视觉处理、短期记忆和语言理解中的作用。在临床方面,科学家也正在研究经颅磁刺激在治疗抑郁症、焦虑障碍和强迫症等方面的潜力。
大脑成像技术
近几十年来,多种脑成像技术的发展极大地提升了科学家在活体状态下观察大脑的能力。
计算机断层扫描通过从多个角度拍摄X射线,再由计算机合成大脑的水平切片图像,提供大脑结构的直观图像,是目前最经济、应用最广泛的结构脑成像技术。正电子发射断层扫描在研究大脑功能方面尤为有价值,通过将放射性标记化学物质引入大脑,追踪血流和代谢活动的变化,从而生成彩色编码的脑功能地图,显示受试者在握拳、歌唱或进行抽象思维时,大脑的哪些区域发生了活跃变化。磁共振成像利用磁场、无线电波和计算机增强技术绘制大脑结构,所提供的图像分辨率显著高于计算机断层扫描,研究表明磁共振成像对于揭示抑郁症患者的海马体萎缩现象具有重要价值。功能性磁共振成像是磁共振成像技术的新变体,通过监测大脑中的血流和氧气消耗来识别高活动区域,具有比正电子发射断层扫描更高的空间分辨率,目前已成为认知神经科学研究的核心工具之一。
脑成像技术的进步为神经科学研究开辟了崭新时代,让科学家得以实时观察活体大脑的工作过程,这在几十年前是难以想象的。但这些技术虽然令人印象深刻,在解读上仍存在相当的局限性和复杂性,需要谨慎对待研究结论。
研究方法的局限性
脑成像技术产生的精美图像,容易给人造成其测量精度远超实际水平的印象。实际上,功能性脑成像只能提供对目标脑区活动状态的粗略近似,而非直接测量神经活动本身。
正电子发射断层扫描和功能性磁共振成像呈现的,是相对于某个基线条件的代谢活动增加区域,“亮起”的区域究竟有多大,在相当程度上取决于选择什么作为比较基线。更重要的是,一个区域代谢活动的增加,并不能证明该区域在特定心理功能中扮演不可替代的核心角色。此外,脑扫描研究需要大量深奥的技术决策,这些决策可能对结果产生实质影响。这些复杂性在一定程度上解释了为何不同实验室的脑扫描研究结果重复性欠佳。尽管如此,这些成像技术仍极大地增强了人类窥探大脑内部世界的能力,是当代神经科学不可或缺的研究手段。
生物学基础的重要性
纵观上述内容,神经元是信息处理的基本单位,通过电化学信号相互通信;不同的神经递质系统分别调节情绪、运动、记忆和认知;大脑的结构层次从后脑的基本生命功能延伸至前脑的复杂思维,各区域各司其职;大脑的可塑性则告诉我们,神经系统并非固化不变,而是能够持续适应和重组。
神经科学与临床实践
现代研究方法从脑电图记录到先进的脑成像技术,为科学家提供了在活体状态下研究大脑的强大工具,使揭示大脑与行为关系的工作取得了前所未有的进展。这些基础研究的成果已经广泛转化为临床实践,中国各大医院神经内科和精神科的诊疗方案,都在不同程度上建立于对神经递质、脑区功能和遗传机制的认识之上。
理解行为的生物学基础不仅帮助我们认识自身,也为神经和精神疾病的治疗提供了科学依据。从帕金森病到抑郁症,从记忆障碍到成瘾问题,生物学研究不断为临床医学开辟新的路径。
可塑性与终身发展
这种生物学视角最具启发性的一点,在于它揭示了人类行为的可塑空间。无论是通过专业训练重塑大脑结构,还是通过改变生活方式影响基因表达,生物学基础并非不可改变的宿命,而是与后天经验持续互动的动态系统。在后续的学习中,我们将继续探索这些生物学基础如何与心理过程、社会环境和文化因素相互交织,共同塑造人类复杂而丰富的行为模式。
大脑的左右分工
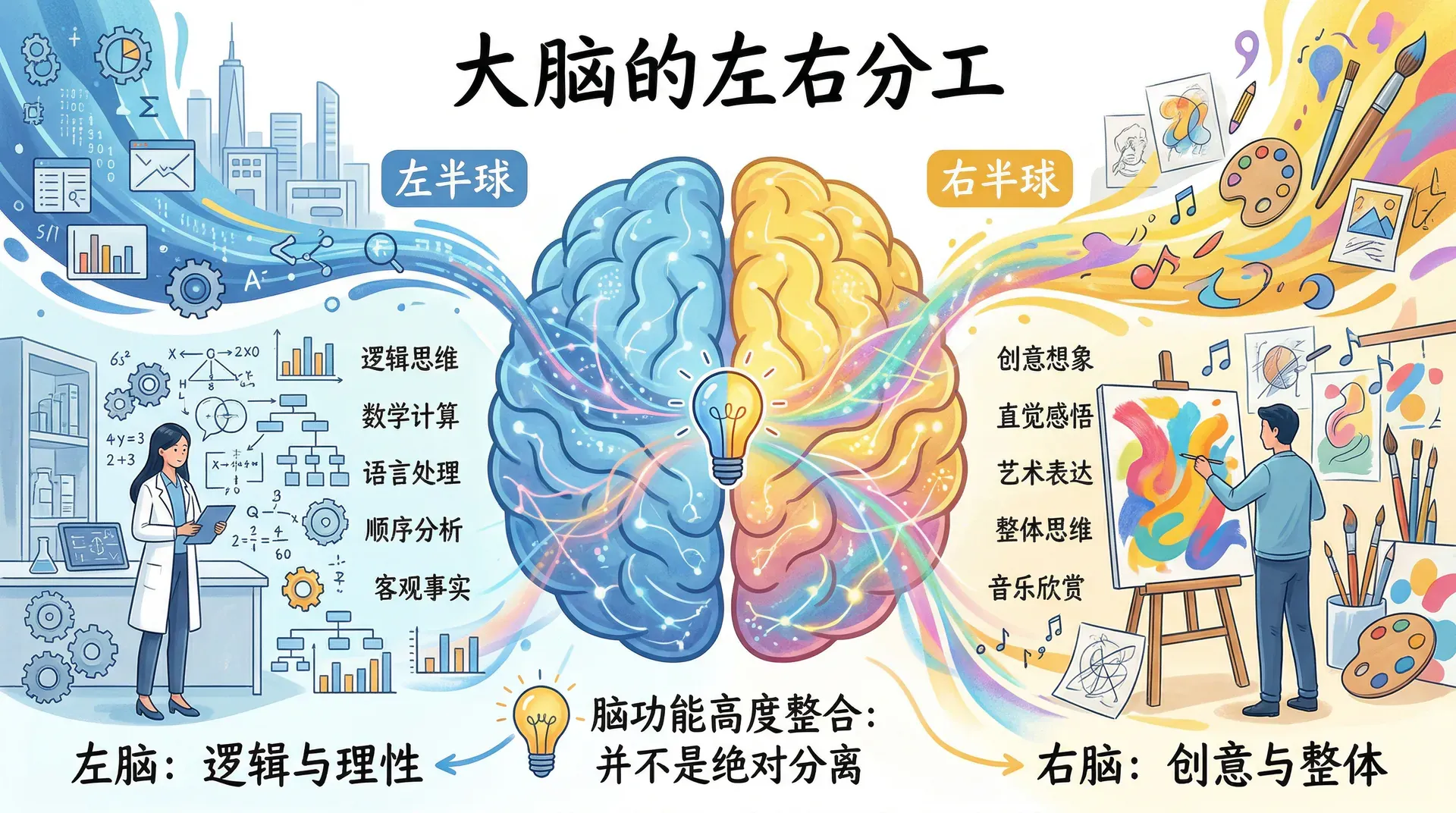
大脑分为左右两个半球,近几十年来,关于两个半球功能专门化的研究非常活跃,由此形成了“左脑逻辑,右脑创意”这一广为流传的说法,尽管这一说法将两者之间复杂的关系大为简化了。
对半球专门化的最早暗示来自脑损伤案例的临床观察。早在1861年,法国外科医生保罗·布洛卡就将左半球与语言控制联系起来。他在治疗一名长达30年无法说话的患者时,发现其言语障碍很可能源于额叶左侧一处局部损伤。此后,大量类似案例证实了这一脑区——后来被命名为“布洛卡区”——在言语产生中的重要作用。1874年,另一个主要语言中心“韦尼克区”在左侧颞叶中被定位,该区域受损通常导致语言理解障碍。由于左半球处理语言的证据日益充分,它一度被称为“主导”半球,而右半球则被视为“非主导”或“哑”半球,这种偏颇认识直到20世纪60年代才开始得到纠正。
揭开半球的秘密
真正改变人们对两侧大脑半球认识的,是罗杰·斯佩里、迈克尔·加扎尼加及其同事对“裂脑”患者的研究。裂脑手术是一种极端的癫痫治疗手段,通过切断连接两个大脑半球的胼胝体,来减少癫痫发作的扩散。这种手术造就了大脑两个半球在功能上实际上被分离的特殊案例,为研究半球功能提供了罕见机会。
理解裂脑研究的逻辑,需要先了解感觉和运动信息是如何路由到两个半球的。左半球控制身体右侧,右半球控制身体左侧。在视觉上,右视野的刺激信息被传送到左半球,左视野的刺激则被传送到右半球。正常情况下,两个半球通过胼胝体实时共享信息,因此人们不会察觉到这种分左分右的“交叉”组织。但在胼胝体被切断后,两个半球之间的信息交流近乎中断,功能专门化因此清晰呈现。
在实验中,研究者将视觉刺激短暂地呈现在单侧视野中,使信息只进入一个半球。当图片呈现于右视野进入左半球时,裂脑患者能流畅地命名和描述所见物体;而当相同图片呈现于左视野进入右半球时,患者无法说出物体名称。这证实了语言功能主要位于左半球。
尽管裂脑患者的右半球无法开口说话,但进一步测试表明它同样在处理信息。当允许患者用左手指认时,右半球能准确识别呈现于左视野的图片,并在视觉空间任务上表现得更为出色。
后续对裂脑患者的研究还发现,右半球在区分颜色、排列积木和识别面孔等视觉空间任务上,往往比左半球表现更佳。这些发现为右半球具有其独特认知优势提供了第一批令人信服的直接证据。
正常大脑中的半球专门化
裂脑研究的一个局限是,切断胼胝体本身制造了一种异常情况,且手术对象都是长期患有严重癫痫的特殊群体,其脑组织在术前可能已有不典型的组织形式。因此,研究者还发展了在普通完整大脑中研究半球专门化的方法。
其中一种方法是测量感知不对称性——即大脑两个半球在处理相同类型刺激时的速度差异。在正常个体中,信息在进入一个半球后会迅速与另一个半球共享,但通过精确测量识别不同类型刺激所需的时间,可以检测出两个半球在处理能力上的细微差异。
总体而言,综合多种研究方法的结果一致表明,左半球在涉及语言处理的任务上更具优势,包括语言理解、阅读和写作;右半球则在许多非语言性任务上表现更佳,包括视觉空间处理、音乐感知,以及辨别他人情绪表情的能力。
半球专门化的复杂性
尽管半球专门化的证据充分,但需要指出的是,一个半球相对于另一个半球的优势在程度上通常相当有限。在正常个体中,两个半球不会单独运作,大多数认知任务都涉及两个半球的参与,且任务越复杂,两个半球协作的可能性就越大。
人们在对两个半球的依赖程度上也存在相当的个体差异。部分人表现出很少的半球专门化,两个半球在各类任务上的能力几乎相当;少数人甚至表现出通常规律的倒转,语言处理可能位于右半球,这种非典型模式在左利手人群中相对更为常见。
有经验的音乐家则提供了另一个有趣的例外。研究发现,经验丰富的音乐家与普通人相比,表现出更多的双侧大脑参与模式,这可能是因为演奏乐器通常要求左右手分别执行不同的精细动作,长期训练促进了两个半球的协同开发。
大脑两个半球的功能专门化是真实存在的,但远比通俗说法描述的要复杂。“左脑理性,右脑感性”的说法将复杂的大脑功能过度简化,现实中两个半球始终是协同工作的整体。
内分泌系统
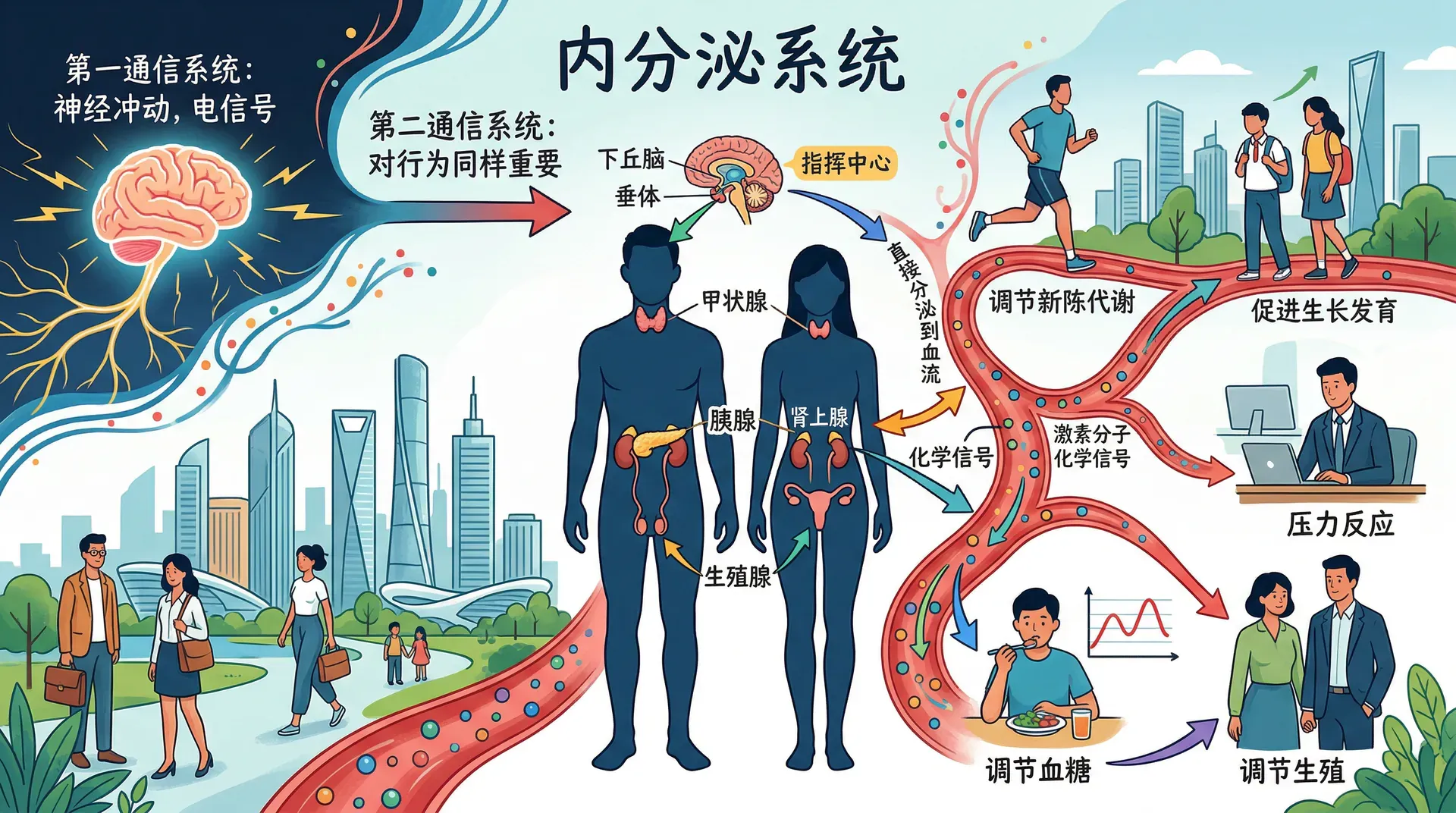
大脑通过神经系统与身体其他部分通信,但身体还拥有第二套对行为同样重要的通信系统:内分泌系统。内分泌系统由一系列腺体组成,这些腺体将化学物质直接分泌到血流中,调控各种身体功能,其信使分子称为激素。
激素在一定程度上类似于神经递质,储存起来以备后续释放,一旦分泌便通过血流扩散,与目标细胞上的特殊受体结合。部分化学物质具有双重身份,在内分泌系统中作为激素,在神经系统中作为神经递质,去甲肾上腺素便是典型案例。激素与神经递质之间存在几个重要差异:神经信号沿着明确的通路以毫秒级的速度传导短距离,而激素信息则通过血流以秒乃至分钟级的速度缓慢扩散至远处,且作用范围更为广泛,可同时影响全身多处目标组织。
激素的释放通常呈脉冲式,一天中在若干时间节点以短暂爆发的形式释放,许多激素的水平在一天之内呈现出有节律的消长变化。内分泌系统的主要腺体包括垂体腺、甲状腺、肾上腺、胰腺和性腺。内分泌系统的大部分功能由神经系统经由下丘脑加以调控,下丘脑与豌豆大小的垂体腺关系密切,垂体腺释放的各种激素扩散至全身,刺激其他内分泌腺体的活动,因此垂体腺有时被称为“主腺”,尽管下丘脑才是幕后的真正调控者。
激素对行为的调节
神经系统与内分泌系统的协同运作,在战斗或逃跑反应中表现得尤为清晰。面临压力情境时,下丘脑同时沿两条路径发出信号——经由自主神经系统和经由垂体腺——到达肾上腺,后者随即分泌“压力激素”,向全身各处扩散,使机体做好应对紧急情况的准备。
催产素是另一种受到广泛研究关注的垂体激素,长期以来以其在女性分娩中触发宫缩和促进泌乳的作用而为人所知。然而,近年的研究揭示,催产素对复杂社会行为同样具有深远影响。实验表明,吸入催产素喷雾能提升个体从细微社会线索中推断他人心理状态的能力,增强对情感情境的移情反应,并在需要信任他人的经济博弈任务中显著提高信任水平,这使催产素获得了“社会黏合剂”的非正式称号。
激素还对人类的生理发育起到关键的调控作用。垂体释放的促性腺激素影响性腺的发育,在出生之前,这些激素指导胎儿外部性器官的形成,使个体的生物学性别在产前发育期间就由激素作用所塑造。青春期性激素水平的大幅升高,负责驱动一系列第二性征的出现,如男性面部毛发和女性乳房的发育;其他激素的变化则触发了青春期左右发生的生长加速。这些由遗传预先编程的激素变化,是理解个体发展中先天与后天关系的重要切入点。
遗传与行为
生物构成影响着个体的行为表现,而遗传构成作为生物学基础的核心,自然也可能对行为产生深远影响。身高、发色、血型等身体特征在很大程度上由遗传决定,这已是广为接受的知识。但智力、情绪稳定性、冲动控制和害羞等心理特征,究竟在多大程度上受到基因的塑造?这些问题是行为遗传学的核心研究领域。行为遗传学是一个研究遗传因素对行为特征影响的跨学科领域,自20世纪70年代以来研究规模持续扩大,为先天与后天的争论提供了越来越丰富的实证依据。颇具讽刺意味的是,行为遗传学在努力证明遗传影响的同时,也不断揭示着环境因素同样不可忽视的作用。
遗传学的基本原理
每个体细胞的细胞核中都携带着来自父母双方的遗传信息,这些信息储存在染色体上,并通过基因加以编码和传递。了解遗传学的基本运作方式,是理解行为遗传学研究成果的必要前提。
染色体和基因
染色体是携带遗传信息的脱氧核糖核酸分子链。人类每个体细胞包含46条染色体,以23对的形式组织,每对中一条来自母亲,一条来自父亲。性细胞精子和卵子各含23条染色体,受精后结合形成拥有46条染色体的受精卵,由此发育出具有完整遗传信息的新生命。
每条染色体又包含数千个称为基因的功能单位。基因是脱氧核糖核酸上充当遗传传递关键单位的特定片段。为什么同一对父母的后代不会是完全相同的克隆体呢?因为性细胞形成过程中,每对染色体的哪个成员进入精子或卵子是随机的,23对染色体可以产生超过800万种不同的组合,精卵结合后能产生大约70万亿种可能的遗传配置。
基因同样成对运作,每对中一个基因来自父方,一个来自母方。当两个基因相同时,该位点处于纯合状态;当两个基因不同时,处于杂合状态。在杂合状态下,对中的一个基因——显性基因——会覆盖另一个隐性基因的表达。显性基因是当配对基因不同时得以表达的基因,隐性基因则在配对基因不同时被掩盖,但仍保留在遗传信息中,可能在后代中重新显现。
基因型与表现型
两个都具有某一显性特征的父母,其后代有时却会表现出该特征的隐性版本,原因正在于基因型与表现型的区别。基因型是指一个人的遗传构成,表现型是指遗传构成在可观察特征上的实际表现。不同的基因型有时可以产生相同的表现型,而同一基因型在不同的环境条件下也可能产生不同的表现型。
基因型在受孕时确定并终身固定,但表现型特征可随时间和环境因素而变化。基因型与表现型之间的关系并非简单的一一对应——并非所有基因对都遵循显性遮蔽隐性的原则,在某些情况下,配对基因不同时会产生介于两者之间的中间表现型。
多基因遗传
大多数人类特征受不止一对基因的影响,这种遗传模式称为多基因遗传。肤色被认为受三到五对基因的相互作用影响,运动协调等复杂身体能力可能涉及更多基因对之间错综复杂的相互作用。心理特征中凡是表现出遗传影响的,几乎都涉及复杂的多基因遗传模式,这也使得精确量化单个基因对行为的贡献变得极为困难。
家庭研究
家庭研究是评估遗传影响的基本方法之一。研究者通过检查血亲在特定特征上的相似程度来评估遗传的作用——如果遗传影响某一特征,血缘关系越近的亲属在该特征上应该越相似。以精神分裂症为例,这种障碍影响约1%的普通人口,但精神分裂症患者的兄弟姐妹中约有9%同样患病,患者的子女约有13%受到影响,而同卵双生子如果一方患病,另一方患病的概率接近50%。这一从普通人群到一级亲属到同卵双生子逐步升高的患病率,支持遗传因素在其中发挥作用的推断。
然而,家庭研究本身无法区分遗传与环境的贡献,因为家庭成员不仅共享基因,还共享相似的成长环境。要更有力地剥离遗传与环境的效应,需要借助双胞胎研究和收养研究。
双胞胎研究
双胞胎研究通过比较同卵双胞胎和异卵双胞胎在特征上的相似程度来评估遗传的影响,其逻辑基础在于两类双胞胎在遗传相关性上的根本差异。同卵双胞胎由一个受精卵分裂而来,拥有完全相同的遗传构成,遗传相关性为100%。异卵双胞胎由两个不同卵子同时受精形成,遗传相关性与普通兄弟姐妹相同,约为50%。由于两类双胞胎通常在相同的家庭环境中成长,若同卵双胞胎在某一特征上表现出比异卵双胞胎更高的相似性,合理的解释是额外的遗传相似性起到了作用。
收养研究
收养研究提供了另一种分离遗传与环境影响的有效手段。通过比较被收养儿童与其生物父母和养父母在特征上的相似程度,研究者可以推断遗传与环境各自的贡献。若被收养者在某特征上更接近从未共同生活的生物父母,遗传因素的作用便得到支持;若更接近养父母,则环境因素的权重更大。收养研究的关键在于,被收养者通常在出生后不久便与生物父母分离,两者之间除了基因之外没有共同的生活经历。
遗传与环境的相互作用
反复的科学研究表明,遗传和环境因素对行为的大多数方面共同发挥影响,且两者之间存在复杂的交互作用,而非简单的叠加。以精神分裂症为例,尽管遗传因素显然参与其中,但似乎没有人会直接“继承”精神分裂症本身,人们继承的更可能是一种程度不等的易感性,这种易感性是否转化为实际的疾病,在很大程度上取决于个人的生活经历和所处环境,某些类型的重大压力事件似乎能在高易感性个体中诱发疾病。
“基因赋予倾向,而不是命运。”这句话精辟地概括了现代行为遗传学研究的核心发现。遗传背景塑造了我们的可能性空间,而具体的人生经历决定了我们在这个空间中落于何处。
近年来,表观遗传学这一新兴领域的发现进一步深化了对遗传与环境关系的理解。表观遗传学研究不涉及脱氧核糖核酸序列改变的基因表达可遗传变化,揭示出特定基因的效应可以通过细胞水平的化学修饰被激活或沉默。更引人注目的是,这些化学修饰可以由环境事件所诱发,包括营养状况、压力暴露或特定生活方式,而且这些表观遗传标记甚至可以传递给后代。这意味着,父辈的生活经历有可能通过生物学机制影响子代的行为特征,遗传与环境之间的边界远比通常所想的更为模糊。
行为的进化基础
要全面理解行为的生物学根源,还需要将视野拉长至进化的时间尺度,思考数百万年的进化历程如何塑造了现代人类和动物的行为模式。进化心理学正是从适应性的角度出发,分析行为背后的进化逻辑,为理解人类行为的共性提供了独特的理论框架。
达尔文的洞察
英国博物学家查尔斯·达尔文并非第一个提出生物进化观念的人,但他在《物种起源》中贡献了对进化变化机制的创造性解释:自然选择。达尔文试图阐明,生物体的特征为何往往恰好适应于其所处的生存环境,以及这种适应性是如何在几代之间逐渐形成的。
在进化理论中,适应性指的是个体相对于种群平均水平的繁殖成功,即留下多少存活后代。某一特征可以通过提供生存优势或繁殖优势来推动进化。自然选择的过程作用于种群,而非个体——当种群基因库的组成因选择压力而逐渐改变时,进化便发生了。这一过程通常极为缓慢,一般需要数千至数百万代才能使一种特征显著普遍化。
提供生存或繁殖优势的遗传特征,比替代性特征更可能传递给后代,因此随时间推移被“选择”下来,这就是自然选择的基本原理。
进化理论的后期完善
达尔文的进化理论在19世纪末至20世纪初遭遇了一个重要质疑:理论缺乏对遗传机制的充分解释,无法说明变异特征究竟如何从亲代传递到子代。格雷戈尔·孟德尔被长期忽视的遗传研究成果在1900年前后重新引起科学界关注,随后数十年间,对遗传机制的深入研究逐渐填补了达尔文理论的这一空缺。到20世纪中叶,将自然选择与遗传学整合起来的现代综合进化论已获得科学界的广泛接受。
当代进化理论的一个核心概念是“适应”,指在种群中因自然选择而增加的遗传特征,因为该特征在出现时有助于解决生存或繁殖问题。由于进化的渐进性,已经不再具有适应优势的特征有时仍会在种群中长期留存。人类对高脂肪食物的强烈偏好便是一个典型案例:在食物匮乏的远古狩猎采集时代,脂肪是稀缺而珍贵的卡路里来源,对脂肪的偏好具有明确的适应价值;然而在食物丰富的现代社会,这一进化遗产却成了肥胖和心血管疾病的风险因素之一。
行为作为适应性特征
物种的典型行为模式常常反映了进化过程中对特定生存挑战的适应性解决方案。研究行为的进化根源比研究形态特征更为困难,因为行为不像骨骼那样能在化石记录中留下直接证据,但通过比较不同物种的行为模式以及分析行为与生态环境的对应关系,可以推断特定行为的进化意义。
中国农村长期观察到一种有趣的现象:野生麻雀在觅食新食物来源时表现出明显的审慎倾向,通常先试探性地少量啄食,等待一段时间确认无虞后才大量进食,而且不会同时尝试两种完全陌生的食物。这种谨慎策略与大鼠等其他动物的觅食行为高度相似,被认为是一种适应性的风险管理行为,可以减少误食有毒食物的概率。
许多行为适应直接指向改善繁殖成功的目标。在众多动物物种中,雌性对交配对象的主动选择是一个普遍现象,雌性通常偏好能提供更好资源、表现出更健壮状态或携带更优质基因的雄性。受青睐的特征往往是雄性健康状况、基因质量或未来服务能力的可靠指标,而非纯粹的随机偏好。
许多看似任意的动物求偶偏好,实际上反映着对潜在配偶遗传质量的深层生物学评估。在人类中,类似的偏好也以更为复杂和文化化的形式存在,这并不意味着人类行为完全由进化决定,但进化的历史确实在人类行为的倾向性中留下了难以抹去的印记。
生物学基础的整合视角
贯穿上述内容,我们从多个层面梳理了行为的生物学基础。神经元作为信息处理的基本单元,通过电化学信号在数以亿计的网络连接中编织出思想与行为;大脑各区域从后脑的基本生命维持到前脑的抽象推理,形成精密的功能分工与协作体系;内分泌系统与神经系统协同运作,以不同的时间尺度和作用范围调控机体的生理与心理状态;遗传与环境的持续交互,从受孕那一刻起就开始塑造每个个体的生物学特质;而亿万年进化历程中自然选择留下的印记,则构成了许多行为倾向的深层底色。
各层面的交织影响
这些层面并非相互独立,而是深度交织的整体。神经递质的水平受基因调控,也受生活经验影响;大脑的结构因学习和训练而重塑;激素的分泌模式随环境压力而变化;进化塑造的行为倾向在文化背景中得到修饰或强化。没有任何单一的生物学因素能够完全解释复杂的人类行为,神经递质系统、大脑结构、激素水平、遗传背景与进化历史,所有因素在相互作用中共同塑造着我们的行为模式。
生物学视角的意义
理解行为的生物学基础不仅帮助我们认识自身的本质,也为神经系统疾病和心理障碍的诊疗提供了坚实的科学依据。这种视角提醒我们:人类行为既有其规律性,也有其可塑性。在后续的学习中,我们将继续探讨生物学基础如何与心理过程、社会结构和文化因素相互作用,共同构筑人类行为的完整图景。