减数分裂与性生命周期
在日常生活中,我们经常可以看到这样的现象:孩子有时候遗传了父亲的深邃眼睛、母亲的高挺鼻梁,甚至还有一些表情、习惯或者性格特点都与父母中的一方相似。然而,孩子并不是父母任何一方的“复制品”,而是一个独特而新颖的个体。比如,两个同胞兄弟姐妹长相不同,有些地方像爸爸,有些地方像妈妈,但又各自具有独立的独特色彩。这种“既像父母又有新变化”的现象,是自然界最巧妙的遗传设计,也是生命演化不断前行的动力。
深入到细胞和分子的层面,从根本上解释这种现象,就离不开一个特殊而复杂的细胞分裂过程——减数分裂。正是通过减数分裂,使得每一代新生命既继承了父母的遗传信息,又在不断的组合与变异中产生了独特的个体变异。这种机制不仅让人类拥有丰富的体貌和性格多样性,也为整个生物界注入了无尽的活力和适应力。
遗传学的基础认知
遗传现象的观察
如果我们观察中国的一个典型家庭,比如著名演员章子怡与女儿醒醒的照片,可以发现女儿可能会遗传到母亲的眼神、父亲的脸型,甚至祖父母的某些特征也可能出现在她身上。有时候,这些特征的组合令人惊叹,仿佛一代又一代生命正以新的排列方式续写着家族历史。这种遗传与变异共存的现象,就是生物学中所指的遗传性状的传递和变异。
除了外貌特征,像血型、身高、疾病易感性等也在家族成员之间代代相传,但每个人又总有自己独一无二的地方。
遗传学是研究生物如何将自身的遗传信息传递给后代,以及这些信息在后代之间为何会出现差异(即变异)的科学分支。它帮助我们科学解释为什么子女既“像”父母,又“有不同”,为我们揭示了生命延续和进化的奥秘。
遗传信息的传递并不是简单的“复制粘贴”过程,而是由一整套精确、复杂的生物学机制来保证的。父母通过各自的生殖细胞,将存储在DNA中的大量遗传信息——可以想象成一套精美的“生命说明书”——传递给子女。这些说明书上的“内容”,决定了我们的外貌、体质甚至某些性格特征。
基因与染色体的关系
要进一步理解遗传的本质,我们就必须了解基因和染色体。基因是遗传信息的最基本单位,就像一条条书写在“生命之书”——DNA上的指令。每个人都拥有成千上万个基因,决定着身高、肤色、血型等各种生物学特征。
这些基因被有序地排列在23对染色体上。可以把人类的全部遗传信息比作一套内容丰富的百科全书,每条染色体就像其中的某一本书,而基因则是每本书中的不同条目。和书一样,每个人的遗传百科全书由父母各贡献一半:一半的染色体来自父亲,一半来自母亲。因此,我们既是父母基因的“拼图”,又在无数基因位点和排列组合中产生了全新的特征。
这样的遗传结构,使得生命的延续既有“稳定性”,也具备了“变化性”。正因为如此,家族成员间既有很多相似之处,又保持着各自的独特性,这也是自然界物种多样性和进化的基础。
繁殖方式的对比分析
无性繁殖的特点
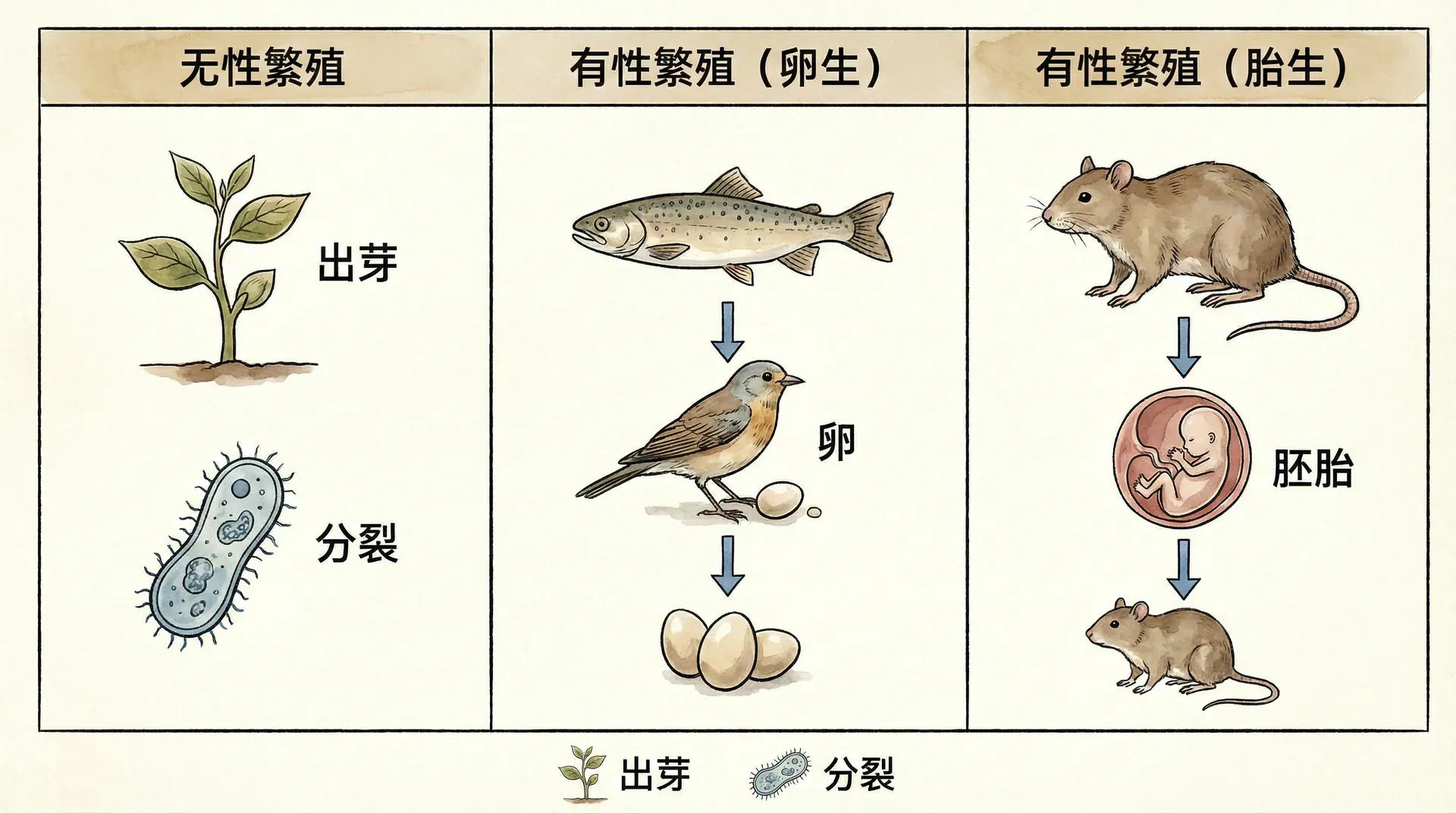
在生物界,繁殖的方式主要分为无性繁殖和有性繁殖。无性繁殖是一种只需要一个亲本,不经配子融合的新个体产生方式。比如我们日常熟悉的马铃薯可以通过块茎繁殖,竹子可以通过地下茎蔓延扩展,这些都是典型的无性繁殖。
无性繁殖有一个显著特点——新个体与亲本在遗传信息上几乎一模一样,就像是“克隆”。中国古代有“ 一母生九子,九子各不同”的说法来形容后代的差异,但无性繁殖下恰恰相反,应该是“九子皆相同”。每一代的新个体都和上一代亲本几乎相同,遗传变异极少。
无性繁殖的优点是繁殖速度快、适用于环境稳定的条件下大量繁殖后代。但缺点同样明显:由于遗传单一,对于环境的变化、突发的疾病或灾害,整个种群的适应能力较弱。只要有一个个体出现遗传缺陷,很可能会影响整个种群的生存。
有性繁殖的意义
与无性繁殖不同,有性繁殖需要两个亲本——一公一母。通过产生和融合含有一半遗传信息的配子(如精子和卵子),形成一个新的生命。虽然这个过程相对复杂,需要耗费更多能量和时间来寻找配偶、产生配子,但由此带来的“遗传多样性”是种群长远发展的巨大优势。
有性繁殖通过基因重组、染色体的自由组合和“交叉互换”等机制,让每一个后代都成为父母基因的全新组合,形成了无数种遗传可能性。正如中国古代名言“物竞天择,适者生存”所强调的那样,遗传多样性让种群能够更好地适应不断变化的自然环境,面对各种疾病、气候变化和生态挑战。
丰富的遗传变异还为进化提供了“原料”,促使物种能够持续优化自身特征。例如,人类对粮食作物、家畜的选育,都是基于这一原理来增强优良性状。
有性繁殖通过减数分裂和受精过程,打破和重组父母的遗传信息,不断创造遗传上独特和多样的新个体。这些个体面对环境时,会表现出不同的适应能力,为物种生存和进化提供了持续的动力与保障。
人类染色体的组织结构
体细胞中的染色体配对
人类的体细胞(也称为“二倍体”细胞,除去精子和卵子等生殖细胞外的所有细胞)中含有46条染色体。这46条染色体实际上可以分为23对,每一对被称作“同源染色体”。在每对同源染色体中,一条来自父亲的精子,另一条则来自母亲的卵子。这种成对的结构是生物遗传的基础。
人类染色体根据结构和功能主要分为两类:第一类是22对常染色体,负责决定绝大部分生理特征和身体发育;第二类是1对性染色体,决定我们的性别。女性的性染色体通常是XX,男性则为XY。
这种成对存在的染色体具有重要的遗传学意义。每对同源染色体上都携带着控制相同类别性状的基因,但具体的等位基因(即基因的不同版本)可能有所差异。例如掌管眼睛颜色的基因,父亲染色体上可能对应“棕色”,母亲遗传的可能是“蓝色”,最终子女的表现型由两者组合决定。这种配对,让我们既承袭父母的共同特性,又具备自己的独特之处。
在细胞分裂期间,同源染色体的排列与分离,决定了每个新细胞所获得的遗传信息组合。而染色体错误的组合或分离异常,也往往是很多遗传疾病发生的根源,比如唐氏综合征就是由21号染色体多了一条所致。
染色体的结构与核型分析
每条染色体本身像极了一本“基因说明书”,包含着大量DNA和蛋白质成分。染色体一般有一个着丝粒,将其分为短臂(p臂)和长臂(q臂),染色体的形态和大小各不相同。通过显微镜观测与核型分析,科学家可以精确判断染色体数量与结构是否正常,这对于疾病诊断、胎儿产前筛查等都具有极高的医学价值。
临床上常见的“核型分析”技术,可以帮助医生发现新生儿是否有先天性染色体异常,如21三体综合征(Down Syndrome),18三体综合征等。这类检测已经成为产前筛查的重要手段。
生殖细胞的形成过程
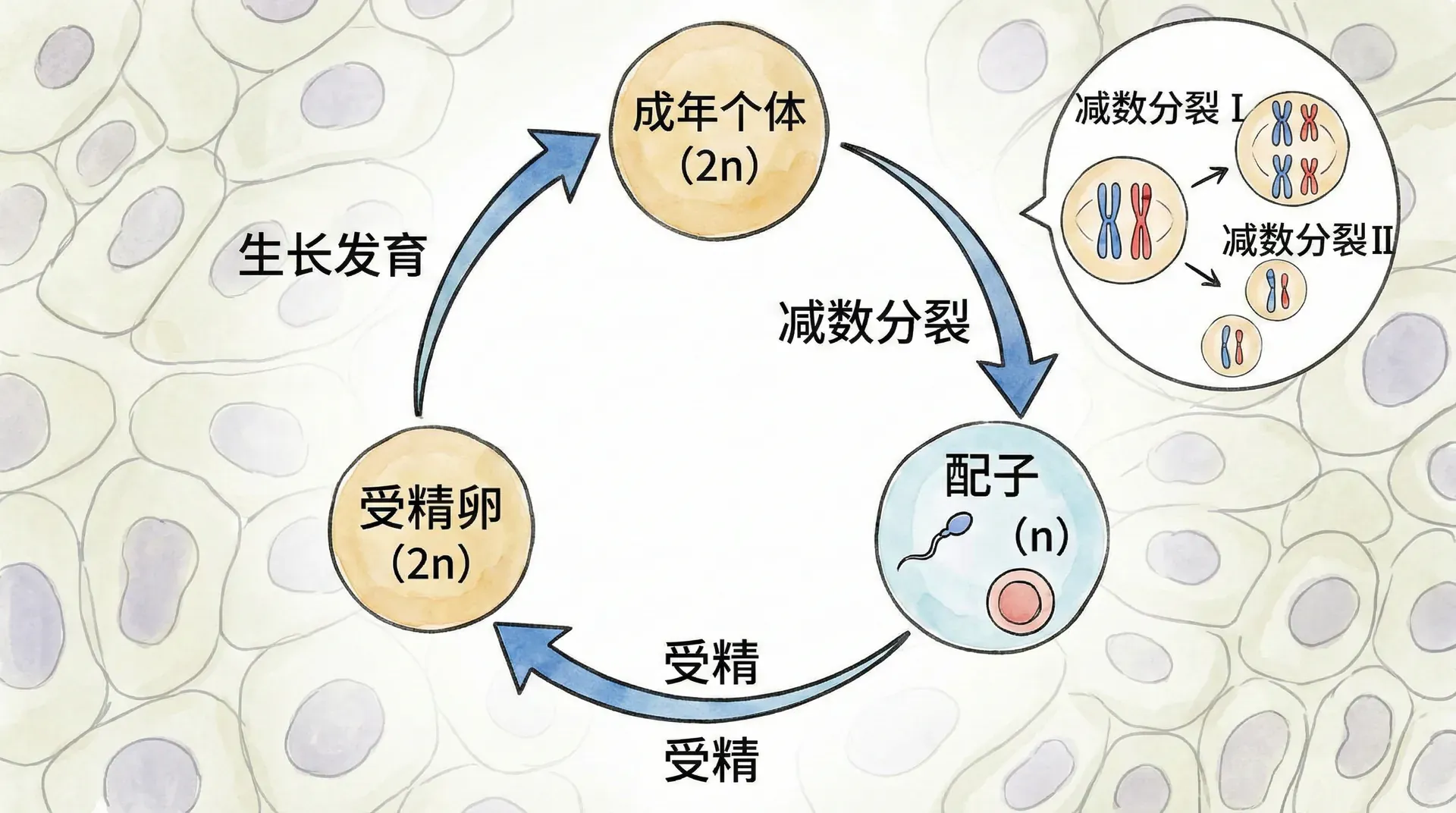
前面提到,人类细胞有46条染色体,但只有体细胞是如此。如果生殖细胞(即精子或卵子)也有46条染色体,那么结合后新个体就会有92条染色体,这显然无法维持遗传信息的稳定和物种的正常发育。
为了解决染色体“翻倍”问题,生物进化出一种巧妙的机制——减数分裂。减数分裂是使生殖细胞染色体数目减半的特殊分裂方式。通过两次连续的细胞分裂,每个生殖母细胞最终会出现4个仅含23条染色体的新细胞(精子或卵子)。这样,受精时来自父母各23条染色体相加,重组为46条,保证每一代的染色体数不变。
减数分裂不仅仅是数量的减少,过程中的“交叉互换”等机制,还极大促进了后代表型与遗传的多样性。这有力推动了人类种群的多样性与适应性。
性染色体的特殊情况
在生殖细胞形成过程中,性染色体(X与Y)也会随机分配。例如父亲产生精子时,有一半精子携带X染色体,另一半携带Y,这就是决定后代性别的根源。如果卵子与Y精子结合则为男孩,与X精子结合则为女孩。
减数分裂的细胞机制
减数分裂第一次分裂(分离同源染色体)
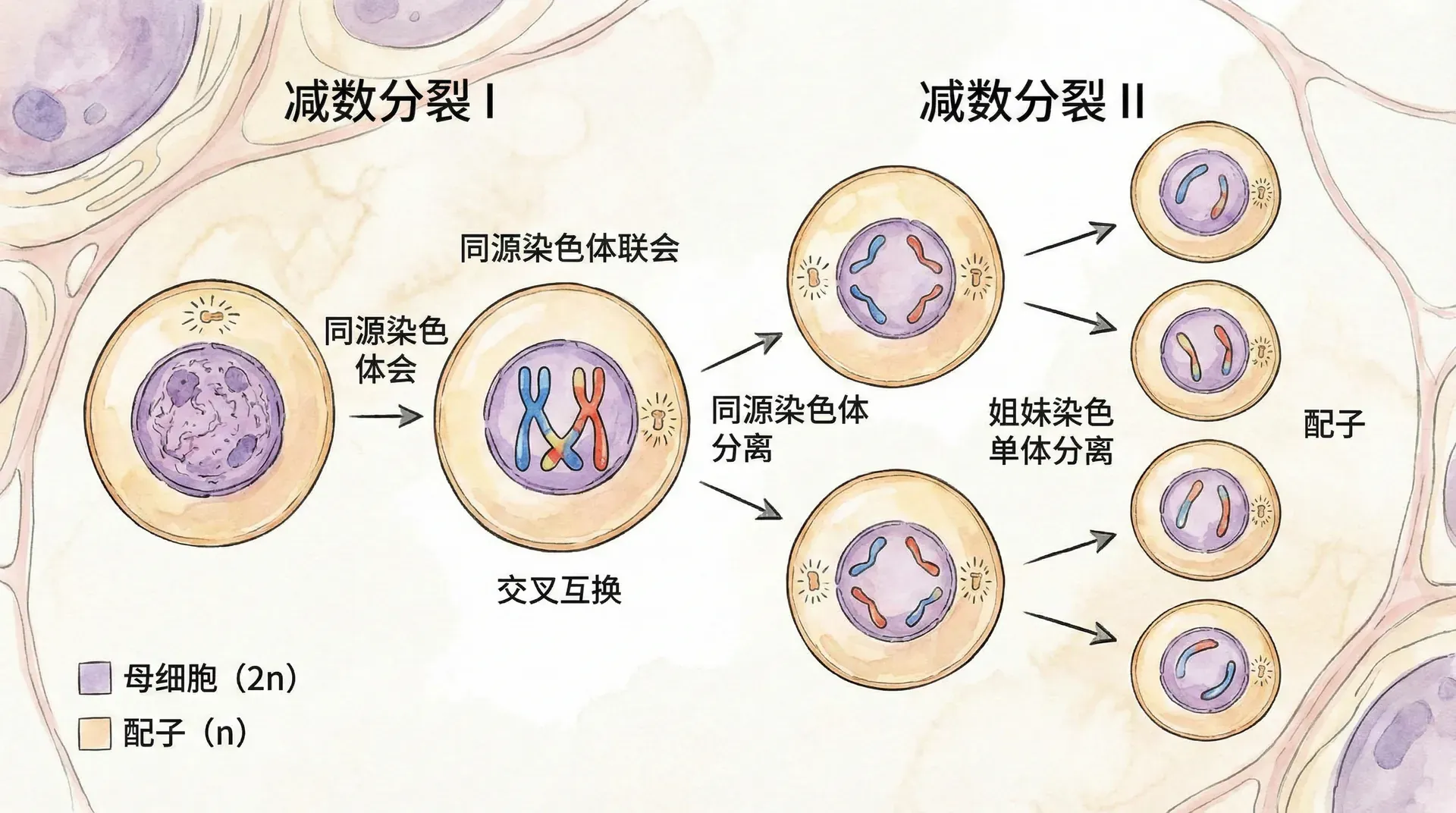
减数分裂分为两大阶段。第一次减数分裂的核心在于“同源染色体分配”——每对同源染色体在细胞的赤道板成对排列,然后被随机分配到不同的子细胞中。这一过程不仅减半了染色体数量,还为基因重组和遗传多样性奠定了基础。
在第一次分裂的前期(前期I),非常关键的“同源染色体配对”和“交叉互换”同步发生。简单来说,这一交换让父母染色体之间的基因片段进行“重新组合”,想象成两份家族食谱互换了一部分内容,使得下一代不再只是简单的“父母复制品”,而有了全新的基因排列。
交叉互换为后代带来了原本不存在的新基因组合,大大增加了遗传的多样性。这一机制正是人类等高等生物能够适应复杂多变环境的遗传基础之一。
第一次分裂完成后,细胞染色体数目由2n降低为n,每条染色体依然以姐妹染色单体的双线状态存在,随后的第二次分裂还会继续精细化基因分配。
减数分裂第二次分裂(分离姐妹染色单体)
第二次减数分裂与普通有丝分裂非常相似,主要作用是将每条染色体的姐妹染色单体进一步分离到不同的子细胞中。最终,一个原始的二倍体生殖母细胞,可以生成四个遗传信息各不相同的单倍体生殖细胞(23条染色体)。
这一完整过程可用数学式表达如下:
如下图所示,细胞中DNA含量会先升高(复制),随后通过两次分裂逐步降低至生殖细胞所需水平:
减数分裂的主要意义有二:一是通过两次连续分裂将染色体减半,确保物种染色体数恒定;二是通过同源染色体的随机分离和交叉互换,极大丰富了后代的遗传多样性,有利于物种进化和适应环境。
与有丝分裂的区别对比和进化意义
减数分裂与有丝分裂虽然在机制上相关,却服务于生物体不同的“人生阶段”:
- 有丝分裂(Mitosis):是体细胞分裂的方式,产生2个与母细胞遗传信息完全相同的新细胞,保证机体生长、修复、再生。
- 减数分裂(Meiosis):专属于生殖细胞的分裂,用以产生带有丰富遗传变异的配子(精子或卵子)。
两个过程各有分工,又协同保障了遗传的“稳定”与“变化”,是生命延续和进化的重要动力。
性生命周期的多样性
动物的生命周期模式
绝大多数动物(包括人类)采用的是“二倍体—单倍体”简单模式:成年个体的体细胞全为二倍体(2n),只有在生殖细胞形成时才进行减数分裂产生单倍体(n)的精子或卵子。受精时,单倍体配子融合,恢复成新个体的二倍体。这种方法保证了属性的稳定传递,也使个体间有了与亲代的变异。
这一生命周期模式让动物可以“稳定繁衍”,维持复杂的器官系统和行为特性。例如,人的体细胞始终维持46条染色体,只有精子和卵子各为23条。
植物的世代交替与生命周期复杂性
比较之下,植物的生命周期就复杂得多了。植物拥有“世代交替”特点,即在其生命周期中,既存在二倍体的“孢子体”阶段,也有单倍体的“配子体”阶段。这两个阶段依次交替进行,实现了遗传物质的循环传递。
例如,我们看到的水稻或玉米植株就是二倍体孢子体。水稻通过减数分裂产生单倍体孢子,这些孢子发育为单倍体配子体(如花粉粒、胚囊),完成配子产生。配子结合产生受精卵(合子),又发育为新的二倍体植株。和动物相比,植物在生命周期上就多了一层复杂和灵活性,也更有利于适应不确定和多变的环境。
特殊生命周期模式
除动植物外,真菌、藻类等生物还存在更多样化的生命周期模式。例如有些藻类主要以单倍体阶段为主,只有在受精时短暂出现二倍体阶段。这些不同的生命周期模式,展示了生物界适应环境和进化的多样性。
人类染色体的精妙组合、减数分裂机制的严密运作,以及动植物各自独特的生命周期模式,共同保证了遗传信息既“守恒”又“变异”,是生命进化与生物多样性的根本动力。
遗传变异的产生机制
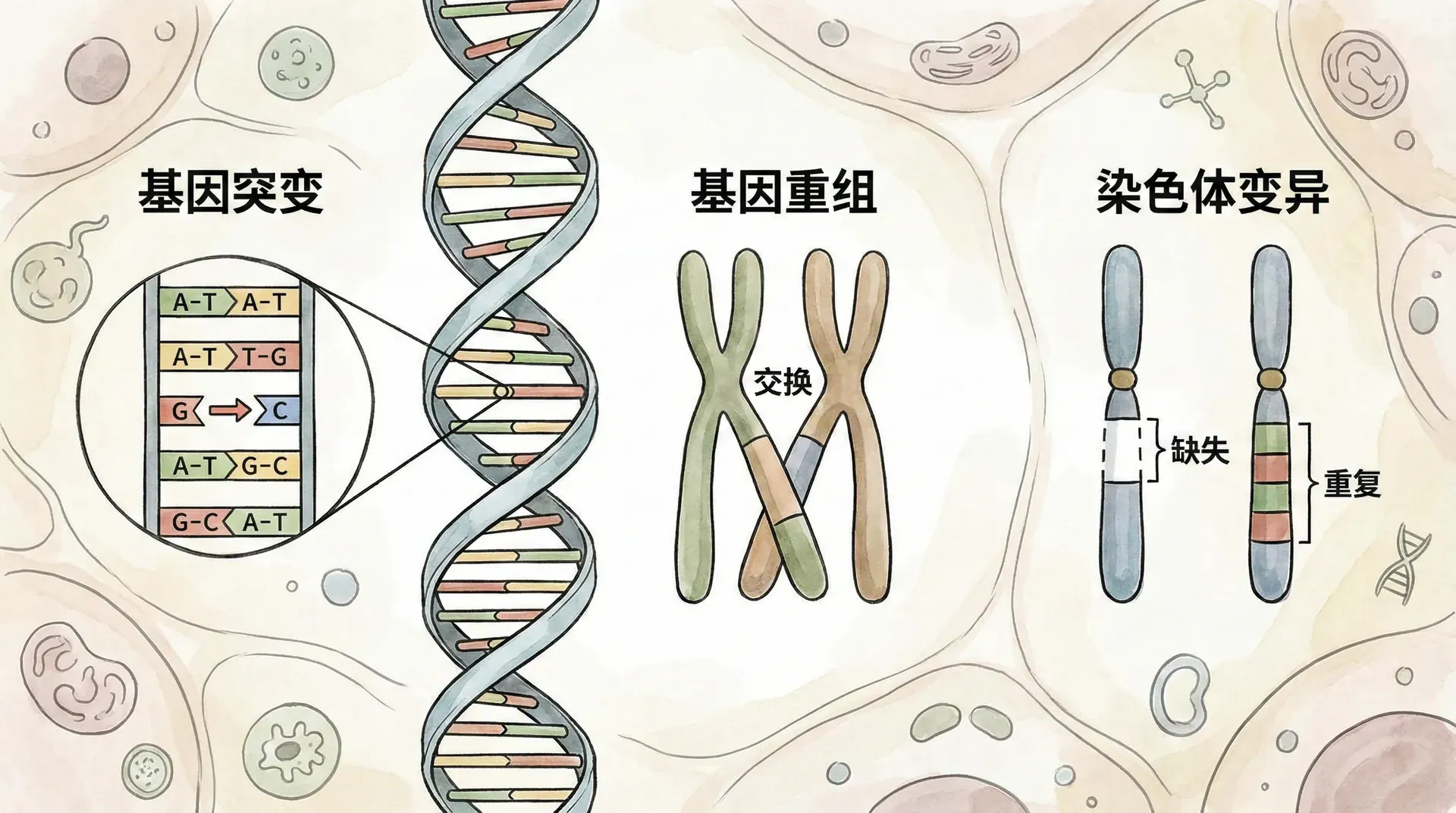
遗传变异是自然界千姿百态的生命现象背后的根本动力。它为生物提供了不断适应新环境、逃避疾病和生存压力的可能性。生物体内的每一项遗传变异,背后都有着极为精巧的机制。主要包括染色体的独立分离、同源染色体的交叉互换、以及受精过程中的配子随机结合。让我们一步步剖析这些机制是如何共同塑造多样化的生命世界的。
染色体的独立分离
在有性生殖中,减数分裂的第一次分裂阶段是遗传变异产生的关键时期。在这个阶段,每一对同源染色体如何分配到子细胞里,完全取决于随机事件。可以想象为“抛硬币”:来自父亲或母亲的染色体进入哪一个新生细胞,并没有先验的规律。这一机制称为同源染色体的独立分离(或自由组合)。
我们有23对同源染色体,在减数分裂时,这23对染色体的分离方式就像同时抛23枚硬币,每一枚都是“正反”两种可能性。这意味着,一个人形成的生殖细胞中,染色体的组合方式有种,也就是大约840万种。这还只是单单考虑染色体整体的组合,并没有算上染色体内部基因的重组!这种巨大的组合数,确保了人群之间的个体多样性。实际上,除非是同卵双胞胎,否则几乎不存在遗传完全相同的两个人。
交叉互换现象
减数分裂不仅通过独立分离带来变异,更重要的还有“交叉互换”现象。所谓交叉互换,是指在减数分裂前期,同源染色体的非姐妹染色单体之间会发生DNA片段的互换。这种基因重组过程,实现了母源和父源染色体间的遗传物质“拼接”,产生全新的基因组合。
在中国人群常见的ABO血型中,一个人如果从父亲那里继承了A型等位基因,从母亲得到B型等位基因,那么由于交叉互换,下一代在生成的配子里可能出现新的组合,甚至有可能产生未有的新基因型。这是为什么同一家族、同一父母所生的子女,在血型、性状甚至疾病易感上都存在差异的根本原因。
据统计,人类每条染色体在减数分裂过程中平均会经历1-3次交叉互换事件。整个基因组以这种方式发生着无数重组,大大提升了遗传变异的种类和概率。
此外,一些动植物会有更高频率的互换,从而赋予种群以极强适应力。例如,某些粮食作物由于长期选育和杂交,染色体交叉互换频率远高于动物,正是这种变异基础,使得我们能够选育出适应不同环境和需求的优良新品种。
随机受精过程
除染色体层面的变异外,受精时精子与卵子的结合同样存在巨大随机性。每一个精子和卵子都包含着不同组合的遗传物质。当这2个分别有840万种可能组合的配子结合时,理论上可以产生大约70万亿种基因型的新个体,这还没有考虑交叉互换和小范围基因突变。
这个天文数字意味着,即使是同一个父母,多次受精产生的子女,遗传背景也是极其独特的。也难怪地球上几乎没有两个完全相同的自然人。
同时,随机受精还有一层重要生物学意义:它进一步稀释了有害基因(隐性致病基因)的影响,提高种群整体的遗传健康度。比如,某些遗传病只有在两个隐性等位基因都遗传给后代时才表现出来,通过随机组合,被“掩盖”的概率大大增加。
遗传变异的进化价值
适应环境变化的意义
遗传变异不仅决定了每一个生命个体的独特性,更是物种整体对环境挑战的“预备队”。如同老子所说:“天下万物生于有,有生于无。”遗传多样性恰如这“有”,为生命演化提供了无限可能。表面上看,变异使同一物种出现了多种多样的表型(如体型、颜色、抗性等),实际上,这些变异就是当环境敲响警钟时,种群内部自我更新与适应的种子。
在近年来全球气候变化中,极端高温、频繁干旱、病虫害暴发等新环境压力不断出现。只有在遗传层面具有高度变异的种群,才能迅速孕育出有利新性状的个体,进而存活下来。例如,中国南方大面积水稻种群中,正是得益于长期自然选择和人工选育形成的丰富遗传多样性,每次旱灾、病害后总有部分品种能幸存下来,并被进一步选育推广。
不仅如此,遗传多样性也有助于生态系统的稳定。当某些性状被环境压力淘汰时,其他性状的个体能顶上,保证物种延续。如同“基因库”的保险机制,变异是整个种群的“免疫系统”。
物种生存的策略
从宏观上看,动物、植物和微生物采用了不同的生存繁殖策略。有性繁殖虽然消耗能量、效率低,但通过染色体独立分离、交叉互换和随机受精,极大地增加了遗传变异,赋予种群强大的环境适应力。而无性繁殖则以速度、复制稳定性见长,适合于短期稳定环境下的爆发式扩张。
可以看到,在环境变化不大、压力较低时(x轴左端),无性繁殖种群由于高效复制反而具备略高的短期适应力。但随着环境变化剧增,只有有性繁殖种群能够依靠强大的遗传变异持续适应,因此在长期生存和进化中占据上风。这正是自然界“动态平衡”的体现。
生物的遗传系统就像一个精密的“风险管理系统”与“创新工厂”:减数分裂和有性繁殖合力制造出千千万万的基因组合,最大限度地为种群预备应对未知的未来。正因如此,地球上的生物能经历一次次灭绝与新生,延续数十亿年的多样生命。
不仅如此,现代生物学对减数分裂和有性生命周期机制的研究,还极大推动了遗传工程、基因疗法、精准医学等高新领域的发展。例如,通过理解并调控交叉互换规律,科学家能够人为增加作物的优良基因重组频率,培育出病虫害抗性更强、产量更高的新品种;又如,通过遗传多样性分析,人类可以提前预防遗传疾病,提高公共健康水平。
从试管婴儿技术、基因诊断,到基因编辑、生产转基因农作物,种种前沿科技的背后,都是减数分裂与遗传多样性理论的坚实支撑。正如中国古代哲学中的“阴阳调和”所揭示的智慧,生命的延续靠的是遗传的稳定传递(守恒)与适应变化(变异)之间的动态平衡。生物通过有性繁殖,将这两者有机统一于一体,成就了无尽演化与辉煌生命。
总之,遗传变异是崭新可能的源泉,也是所有生物跨越险境、繁荣昌盛之本。深入探究和把握这些机制,不仅让我们领悟自然界的奥妙,还能为人类自身的发展和福祉开辟更为广阔的前景。