生物多样性与群落稳定性
生态系统中到底能够容纳多少种生物?一个区域内的物种数目的上限是多少?是什么决定了这些差异?比如,为什么热带雨林中生物多得数不过来,而寒冷的北极苔原里却只有寥寥几百种?这些看似简单的问题,其实隐藏着生态学中关于物种多样性与群落稳定性的核心谜题。不同的地区、气候、资源供给和历史事件都深刻影响着物种的分布和数量。
科学家们一直在尝试解释这些差异,探索生物多样性背后的规律。今天,我们就将一探究竟:是什么限制了生态系统能容纳的物种数量?是什么机制让一个系统保持稳定,又是什么让某些系统更加脆弱?通过深入了解,我们也可以思考人类活动又如何影响着这些生态系统的平衡。
生态系统的饱和现象
个体数量的饱和
在一个封闭的生态系统中,植物通过光合作用固定太阳能,这些能量随后被食草动物、肉食动物和分解者逐级利用。当系统达到平衡时,所有生产出来的能量都会被消耗掉,这时我们说这个系统在个体数量上达到了“饱和”状态。
以长江中下游的湖泊生态系统为例,湖水中的藻类每天产生一定量的有机物,浮游动物、小鱼、大鱼依次捕食,最终能量在整个食物网中流转完毕。此时,湖泊已经容纳了它所能支持的最大生物量。
生态位是指一个物种在生态系统中的位置和作用,包括它在哪里生活、吃什么、什么时候活动等所有生态特征的总和。就像每个人在社会中有自己的职业和生活方式一样,每个物种也有自己独特的生存方式。
但实际情况往往更复杂。捕食者通过捕食会降低猎物的数量,使猎物种群无法达到理论上的最大密度。在青藏高原的草地上,狼群的存在限制了藏羚羊的数量,使得草地从未真正被完全啃食。只有处于食物链顶端的捕食者,才可能达到相对完全的饱和状态。
物种数量的饱和
更有趣的问题是:一个生态系统能容纳多少个不同的物种?这个问题的答案关系到生态系统是否存在“物种饱和”现象。如果一个生态系统已经物种饱和,那么新物种要么无法成功入侵,要么必须导致一个现有物种灭绝才能站稳脚跟。反之,如果新物种能够成功定居而不引起其他物种消失,说明原来的系统还没有达到饱和。
一项经典的对比研究提供了物种饱和的有力证据。观察发现,在中国、北美和澳大利亚,植被结构相似的栖息地中,鸟类物种数量惊人地相近。下图展示了这种关系:
植被垂直结构越复杂(从地面草本层到低矮灌木层再到高大乔木层),鸟类物种数就越多。关键在于,植被分了多少层比具体是什么植物更重要。一个拥有草地、灌丛和高大树木的森林,能支持的鸟类物种数量,远超单一结构的草原。
这种现象暗示着,鸟类群落可能已经达到了物种饱和状态。但对于植物、昆虫和蜥蜴等其他生物类群,类似的规律并不总是成立,这说明不同类群的饱和程度可能存在差异。
物种多样性的奥秘
多样性的度量
走进云南西双版纳的热带雨林,你随手指向一棵树,很难猜出它是什么种类,因为这里有数百种树木混杂生长。但在华北平原的玉米地里,你闭着眼睛都知道脚下是什么植物。这就是物种多样性的直观差异。
物种多样性不仅包括物种数量(物种丰富度),还包括各物种的相对重要性。两个生态系统可能都有50个物种,但一个是少数几种占据主导地位,其余都很稀少;另一个则是各物种数量相对均衡。后者的多样性更高。
下图展示了三种不同的物种重要性分布模式:
长白山原始混交林中,各树种的数量较为接近,没有绝对的优势种。而在干旱的塔克拉玛干沙漠边缘,红柳和梭梭占据压倒性优势,其他植物只能在缝隙中艰难生存。
多样性的三个维度
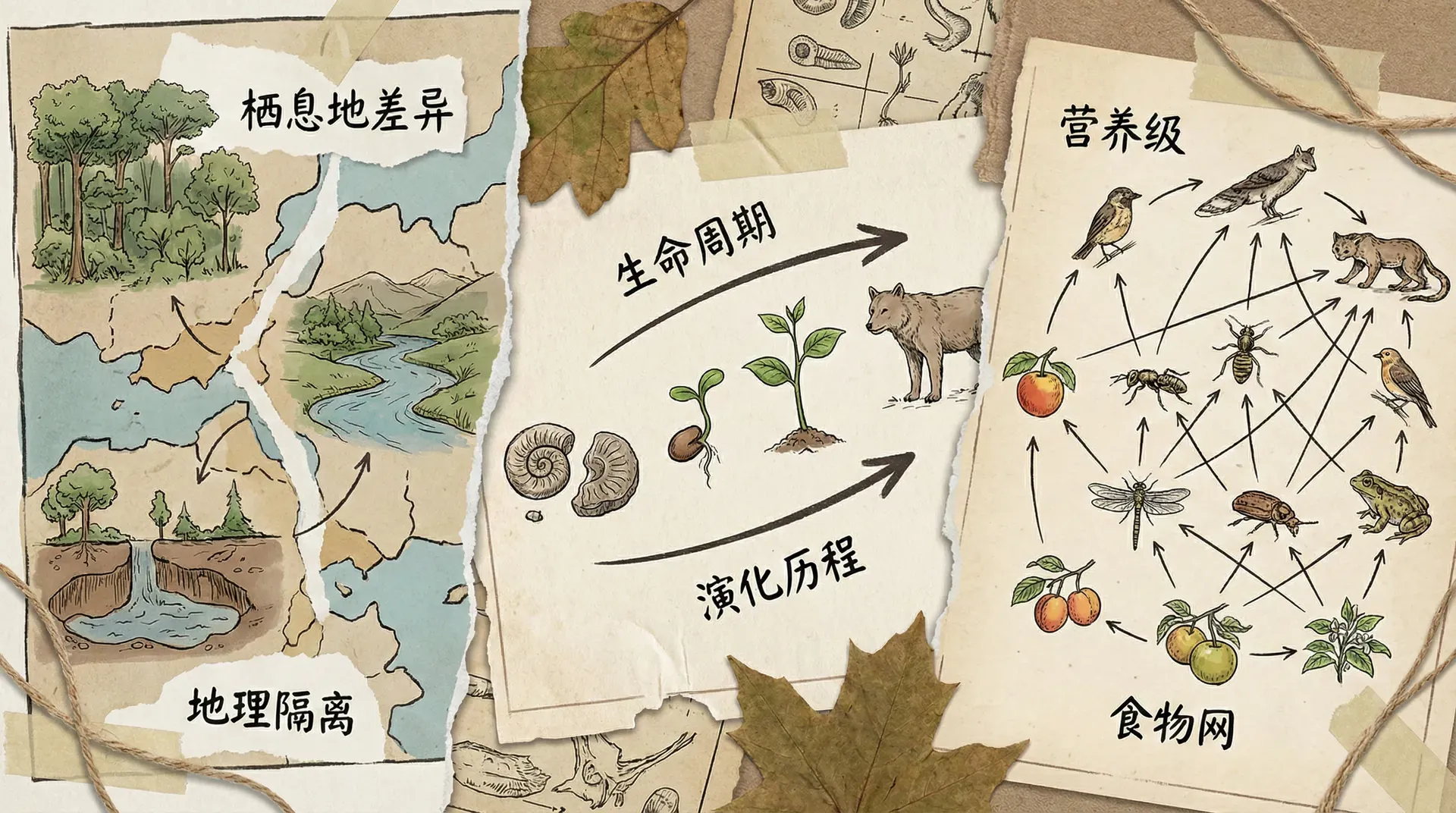
生态系统的物种多样性可以从三个基本维度来理解:空间、时间和食物。
空间维度
不同物种在水平和垂直方向上分工明确。在一片森林中,有些鸟类专门在树冠层觅食,有些在树干上攀爬寻找昆虫,还有些在林下灌丛中穿梭。同样,在长江口的滩涂上,不同种类的鹬在不同深度的泥层中寻找食物,各自占据特定的觅食位置。
以西双版纳热带雨林为例,植被从地面到树冠可以分为地被层(0-0.6米)、灌木层(0.6-8米)和乔木层(8米以上)。这种垂直分层为不同生态习性的动物提供了多样的栖息环境。在林冠层活动的白颊长臂猿几乎不会与地面的野猪发生资源竞争。
时间维度
物种还可以通过时间上的分离来共存。在神农架的森林里,猫头鹰在夜间捕鼠,而日行性的猛禽在白天活动,两者虽然捕食相似的猎物,但通过时间错开避免了直接竞争。
季节性的分离同样重要。青藏高原的高山草甸在不同季节开放不同的花朵:早春的雪莲,仲夏的龙胆,晚秋的绿绒蒿。相应地,传粉昆虫的种类也随季节更替,形成了时间上的物种替换。
营养维度
食物选择的差异是物种共存的重要机制。即使生活在同一片水域,不同鱼类也有各自的食谱。长江中的草鱼吃水草,鲢鱼滤食浮游植物,鳙鱼吞食浮游动物,青鱼则专吃螺蛳。这种食性分化让多个物种能够共享同一片水域。
体型差异通常对应着食物大小的差异。在同一片湿地,小型鹭鸟捕捉小鱼小虾,中型的苍鹭捕食较大的鱼类,而大型的白鹤则能处理更大的猎物。这种分级利用资源的方式,极大地提升了群落的承载能力。
生物多样性的三个维度——空间、时间和食物——相互交织,共同决定了一个生态系统能够容纳多少物种。理解这三个维度,是保护生物多样性的基础。
从赤道到两极的多样性变化
纬度梯度现象
地球上存在一个引人注目的规律:从赤道向两极,生物多样性逐渐降低。海南岛的热带雨林可能在一公顷内就有上百种树木,而黑龙江的针叶林往往只有寥寥数种。
让我们看看中国境内爬行动物多样性随纬度的变化:
位于北纬20度左右的海南岛,蜥蜴种类最为丰富;随着纬度升高,到了东北地区,蜥蜴几乎绝迹。这种普遍存在的纬度梯度背后,究竟是什么在起作用?
影响多样性的机制
生态学家提出了多种机制来解释物种多样性的差异。这些机制可以分为不同层级,有的直接受物理环境影响,有的则涉及生物之间的相互作用:
让我们用几个例子来理解这些机制:
气候稳定性与可预测性
云南南部的气温年变化幅度只有10℃左右,许多昆虫和植物可以高度专化——有的植物只在特定月份开花,有的昆虫只取食某一种植物。相比之下,内蒙古草原的年温差超过50℃,这里的生物必须具备广泛的适应能力才能生存,因此专化程度低,物种数量也相对较少。
生产力假说
这个机制特别有趣。当食物充足时,动物可以挑食。例如,一片沙漠绿洲植物稀疏,蜥蜴见到昆虫就必须捕食;另一片植物茂盛,昆虫丰富,蜥蜴可以专门挑选最容易消化、营养最好的种类。结果是,后者能养活更多种对食物有不同偏好的蜥蜴。
图中展示了不同生态系统的资源可用量,高生产力环境的资源是低生产力环境的两倍。彩色矩形代表各个物种的生态位宽度。可以看到,在资源充足的环境中,每个物种可以占用更窄的生态位(更专业化),因此相同范围的资源能支持更多物种。
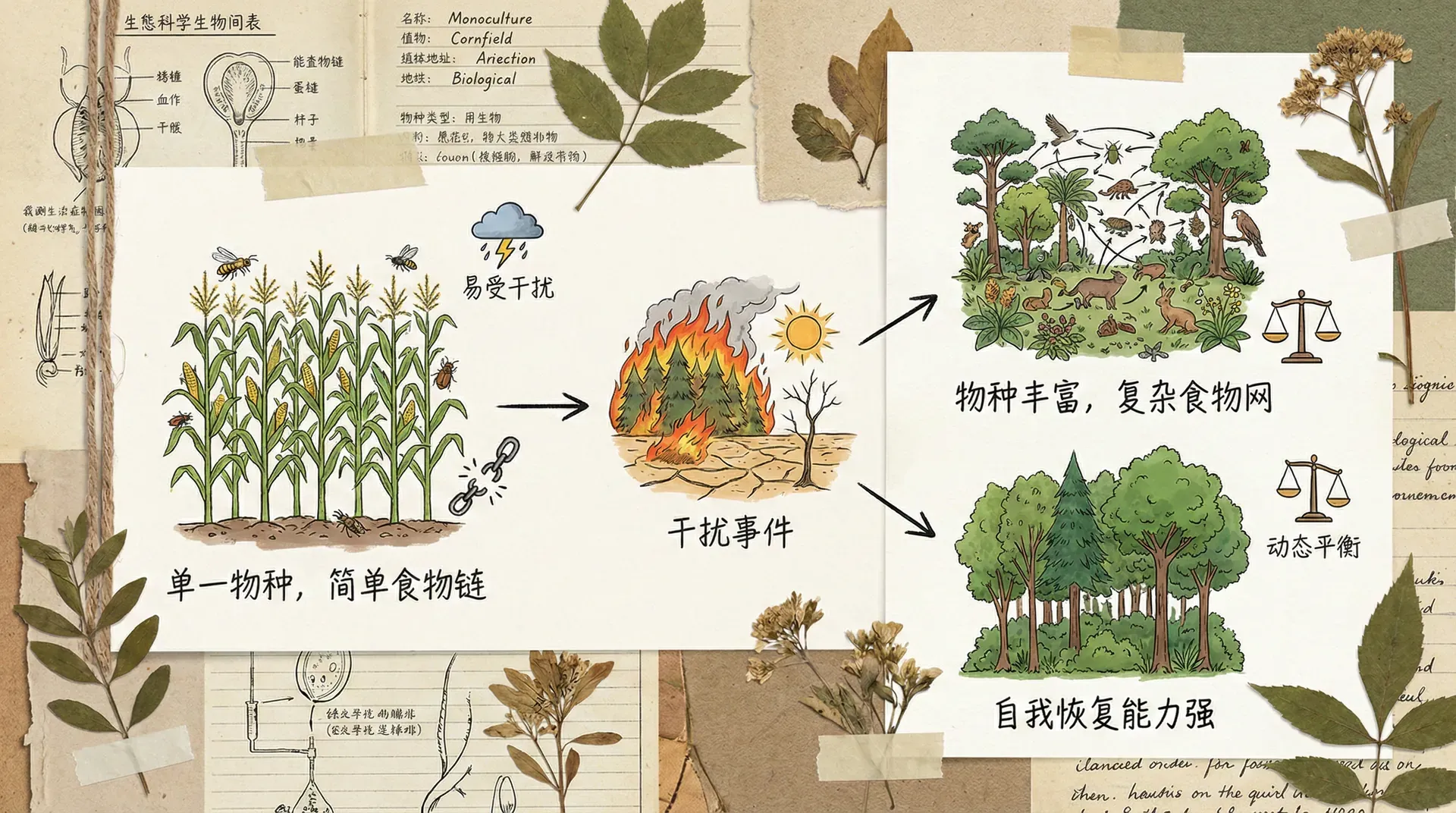
适度干扰假说
这个假说特别能解释一些令人费解的现象。极端稳定和极端多变的环境,多样性都不高;而处于中间状态的环境,多样性反而最高。
长江中下游的洪泛平原就是很好的例子。每年夏季的洪水会淹没部分区域,但洪水是可预期的,强度也适中。这种周期性的干扰阻止了某些竞争能力强的物种完全占领空间,为生长较慢但更耐淹的物种保留了立足之地。如果完全没有洪水,几种强势植物会排挤掉其他物种;如果洪水过于猛烈频繁,大部分物种都无法生存。
生态系统的稳定性
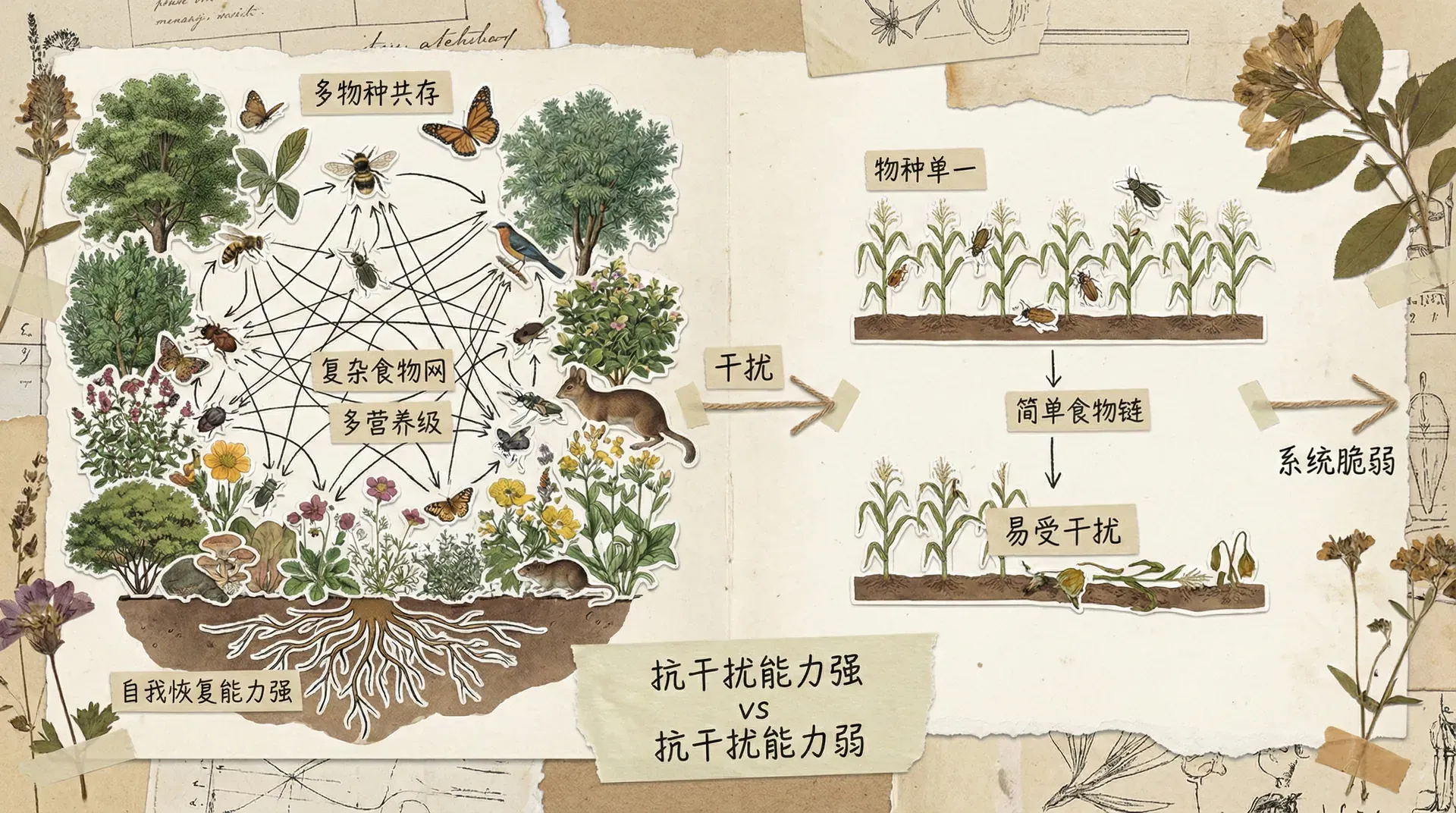
稳定性的多副面孔
当我们说一个生态系统“稳定”时,实际上可能指完全不同的意思。稳定性至少包含四种不同的含义,而一个系统可能在某种意义上很稳定,在另一种意义上却很脆弱。
多样性与稳定性的关系
一个古老的生态学观点认为:物种越多,生态系统越稳定。逻辑似乎很简单——物种多意味着食物网复杂,能量流动途径多样,某个物种波动的影响可以被其他途径缓冲。
但理论分析给出了出人意料的结果。数学模型显示,随意组装的复杂系统(物种多、相互作用强)反而更容易失稳。这是因为复杂的相互作用网络中,一个物种的扰动会通过多条路径传递和放大,最终导致整个系统振荡或崩溃。
这个悖论困扰了生态学家数十年:现实中的复杂生态系统明明长期存在,为什么理论却说它们应该不稳定?答案可能在于,自然系统的构建遵循特定规则,而不是随机组装的。
实地观察的结果并不一致。有研究发现,草原植物种类越多,抵御干旱的能力越强,干旱过后恢复也越快。2000年以来,黄土高原大规模退耕还林还草,植被多样性增加,水土流失减轻,生态系统稳定性明显提升。
但也有相反的例子。加拿大森林中,以单一针叶树为食的昆虫,种群波动反而比食性广泛的昆虫更小。这可能是因为专食性昆虫与寄主植物之间形成了更精细的调控机制。
目前的共识是:多样性与稳定性的关系取决于具体情境。不同类型的稳定性对多样性的依赖程度不同,不同的生态系统和生物类群也表现出不同的模式。但总体而言,维持较高的生物多样性,是生态系统健康和可持续性的重要保障。
群落动态的数学描述
生态学家用数学方程来描述物种之间的相互作用。最基础的模型可以写成:
这个方程表示第 (i) 个物种的数量 (N_i) 随时间的变化率。其中 (b_i) 是该物种在没有其他物种影响时的增长率,α下的求和符号代表第 (j) 个物种对第 (i) 个物种的影响强度,可正可负。如果是负值,说明存在竞争或捕食;如果是正值,则可能是互利共生。
这个看似简单的方程,实际上包含了丰富的生态学含义。当系统包含多个物种时,它们的命运相互交织。一个物种数量的增加,可能通过竞争压制另一个物种,但又可能为第三个捕食者提供更多食物,而这个捕食者的增多又反过来控制第一个物种……
系统的稳定性取决于这些相互作用的强度和结构。如果物种自我调节能力强(数量增加时受到负反馈强烈),相互作用又比较温和,系统就容易保持稳定。但如果相互作用很强,物种数量又很多,系统就可能出现复杂的振荡甚至混沌行为。
真实的生态系统之所以能够稳定存在,是因为长期演化筛选出了特定的相互作用结构。不是所有的物种组合都能共存,只有那些相互作用符合特定约束的组合,才能形成持久的群落。
保护启示
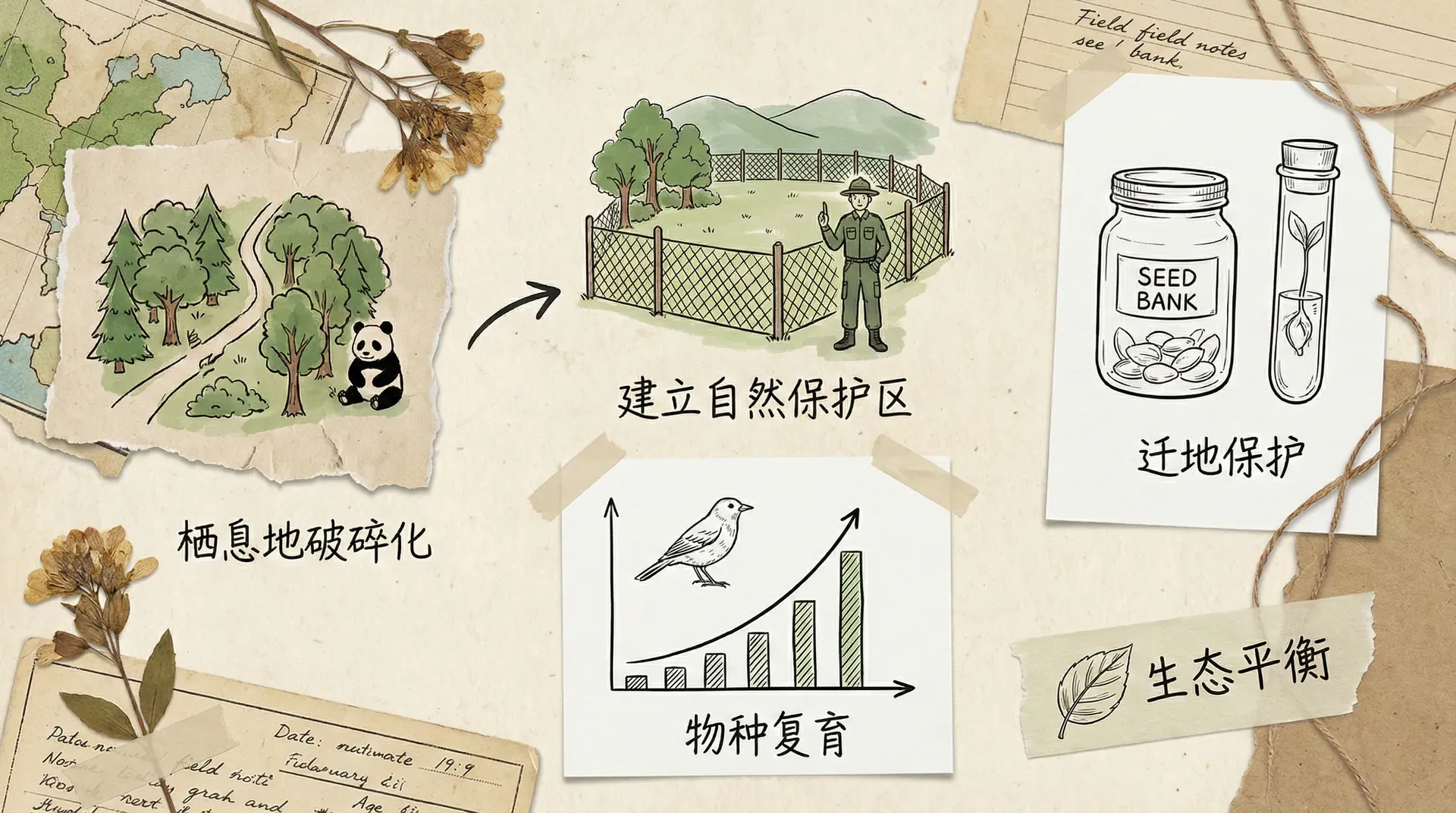
理解生物多样性的形成与维持机制,对于生态保护具有重要意义。
-
保护需要考虑空间尺度。一个小的保护区可能只能保护点位多样性,但区域多样性需要保护足够大的面积,包含不同的生境类型,让物种能在空间上更替。
-
适度的干扰并非坏事。完全封禁可能让某些强势物种占据主导,反而降低多样性。在三江源保护区,允许适度放牧的区域,植物多样性有时反而高于完全禁牧区。
-
保护关键种和关键过程至关重要。顶级捕食者的消失会引发级联效应,改变整个食物网的结构。同样,传粉昆虫、种子传播者等功能类群的丧失,会破坏生态系统的正常运转。
-
多样性不仅关乎物种数量,更关乎生态系统的功能和服务。一个功能完整、相互作用网络健全的生态系统,才能真正具备抵御环境变化、持续为人类提供服务的能力。
从西双版纳的热带雨林到青藏高原的高寒草甸,从长江三角洲的湿地到大兴安岭的针叶林,中国辽阔的土地上孕育着丰富多样的生态系统。保护这些生态系统的多样性,不仅是为了自然本身,更是为了我们自己和子孙后代的福祉。