生物进化与自然选择

生物进化是贯穿整个生物界的核心过程,无论是在细胞内部基因组的一丝微小变异,还是在全球尺度上生态系统结构的更替,都能见到进化的印记。基因突变、染色体重组以及选择压力共同作用,使得生命不仅在形态外观上千变万化,更在行为和生理机制上体现巧妙适应。例如,在中国南北跨度和环境类型极为丰富的背景下,许多动植物通过调整繁殖策略、能量代谢或捕食方式,逐步适应从湿润低地热带到寒冷高原荒漠等多样的生态环境。
这些区域性的适应和多样性演化,不仅反映出生命对自然挑战的应对能力,也展现了中国生物多样性呈现多层次、多样化格局的形成原因。透过观察这些不断变化的生命现象,我们能更深刻地理解进化如何塑造和维护着地球生态的复杂与美丽。
进化的驱动力量
当我们谈到生物进化时,实际上指的是群体中基因频率随时间发生的改变。在自然界中,个体虽然表现出变异和适应性,但真正意义上的进化其实是群体层面的基因库在不断变化。推动生物进化的力量主要包括:基因漂变、基因流动、减数分裂偏差、基因突变以及自然选择。
在这五种进化力量中,只有自然选择是有方向性的,会推动有利于生存与繁殖的基因在群体中增加,使生物更好地适应特定环境。其余几种力量,如基因漂变、基因流动和突变,则更多地为群体带来新的变异或随机变动,有时甚至会导致适应性变差。
这些进化驱动力不仅单独作用,还可能共同影响群体的遗传结构。在自然环境、地理隔离、气候变迁等多重因素影响下,不同地区和物种的进化轨迹呈现出极为丰富的多样性。我们可以通过野外调查、分子生物学实验和模拟模型等多种方法,对这些力量如何影响自然界中的生命演化进行深入研究和理解。
自然选择的三种类型

自然选择通过筛选不同性状的个体,塑造群体的遗传结构。根据作用的方式和结果,我们通常将自然选择分为三种主要类型:稳定化选择、定向选择和分裂选择。这些模式不仅决定了群体的遗传多样性,还影响着物种如何适应环境的变化。下面我们通过具体实例详细说明。
稳定化选择
当环境条件相对长期稳定时,群体中表现中等的或“平均型”个体通常拥有最高的生存和繁殖成功率。这种选择会“压制”极端性状,使中间型性状更为常见,从而减小个体间的差异,维持群体的遗传稳定。例如,以中国人的身高分布为例,过高或过矮的人在历史上的生存挑战会更大,如更容易受到外伤、能量利用效率不理想等;而身高适中的人则可能在体力、能耗以及社会交往等方面更具整体优势,因此更容易留下后代。
这种现象在自然界中十分普遍,比如大多数鸟类孵化出的卵大小适中存活率最高;新生鼠的体重过大或过小都不易成活。稳定化选择有助于排除极端变异,长期来看,会使群体表现出较窄的性状分布范围。
定向选择
定向选择发生在环境因素发生变化或迁入新环境时。当原有的最适性状不再适合,新的极端性状开始受到青睐,这时群体性状平均值会向某一方向“移动”。工业革命后的“工业黑化”现象是著名案例:在英国,由于空气污染,树皮颜色加深,深色的斑蛾在工业区更易伪装和生存,于是深色型个体显著增加。而在中国东北和华东某些工业化城市,也发现了类似现象:传统浅色昆虫因对污染环境的暴露而易被天敌发现,深色个体生存优势明显上升。
这种选择会导致群体遗传特征发生快速改变,比如农作物在持续气候变暖的情况下,逐渐表现出耐旱和耐热的特性;高原动物逐步演化出更高的红细胞数量应对低氧环境。定向选择是群体适应新环境的重要动力。
分裂选择
分裂选择,又称“分化选择”或“两极选择”,指的是中间型个体在环境中反而不如两种极端型个体有利。这通常发生于环境出现显著多样性、不连续变化时。中国一些蝴蝶的拟态现象是经典例证:部分蝴蝶翅膀颜色要么接近绿色叶片,要么像极枯黄落叶,以逃避天敌捕食,而翅色中等的个体更容易被发现、捕食。这样一来,群体里极端色型的比例增加,而中间型被"淘汰"。
除了蝴蝶,中国西南地区生活于不同海拔的小型蛙类,有的体色适应湿润的苔藓,有的适应干燥的褐色泥土,中间型个体因无法有效伪装而存活率较低。分裂选择有助于推动群体内产生更丰富的多样性,甚至可成为新物种形成的开端。
频率依赖选择
除了上面三种经典模式外,自然选择还可能随性状在群体中的频率而变化,这就是频率依赖选择(frequency-dependent selection)。在其中,个体适应度受其性状在整体中的分布影响:有时罕见的性状反而有利(负频率依赖);有时流行性状更容易取得优势(正频率依赖),但在自然界,负频率依赖选择更为常见且对生物多样性有重要意义。
比如,某些鱼类或昆虫存在不同的色型或骗术型,天敌往往优先攻击最常见的类型,使得罕见类型暂时变得安全,比例又会回升。如此循环往复,保证了多样性的维持。
负频率依赖选择是维持生物多样性的重要机制。当一种性状变得稀少时,它往往能获得优势,从而防止该性状完全消失。
生态遗传学实例
我们来看中国南方地区的华南虎纹蛱蝶,它们是一种具有显著多态性的蝴蝶。在不同季节,森林环境背景的显著变化,对不同翅膀颜色的个体施加不同的选择压力。
在春季和早春,地面多为褐色干叶与树枝,此时翅膀为褐色的个体易于隐蔽,被鸟类天敌捕食的概率较低。进入夏季随着树林茂密、绿色覆盖,大量绿叶的出现使得绿色翅膀的蛱蝶能够更好地融入环境,存活率大幅提升。而此时,褐色翅蝶则变得显眼,捕食率上升。
这一现象不仅体现了季节性选择的作用,也表现出负频率依赖:当某一色型逐渐变多,天敌更易辨识并产生“学习效应”,导致该型捕食率升高。这样,两种翅色的个体数量随着季节和捕食压力交替领先,避免了任何一种色型的完全消失,从而维持种群的多样性与生态的平衡。这种机制在其他生态系统和生物中也相当常见。
这种季节性的选择压力变化正是负频率依赖选择在起作用,确保了两种色型在群体中都能保持一定的比例,体现了自然界维持遗传多样性的智慧。这也为我们理解物种多态性和生态平衡提供了生动的实例。
新物种是如何产生的
物种形成是进化生物学中最引人入胜、最核心的问题之一。理解新物种产生的过程,有助于我们深入认识生物多样性的根本来源。总体来说,新物种的形成主要包括两种经典途径:异域物种形成和同域物种形成,但除此之外还有一些更为复杂、交织的特殊情形。
异域物种形成
异域物种形成(allopatric speciation)是由于地理隔离导致的物种形成模式。当一个物种的分布区被江河、山脉、戈壁等自然屏障分隔开,原本可以自由交配的群体被分割成多个小群体。随着时间推移,不同区域的环境条件和自然选择压力各异,同时基因漂变和突变也持续累积,最终这些群体的基因组发生了不可逆转的差异。
中国幅员辽阔、地貌复杂,拥有许多天然的“自然实验室”。例如,青藏高原的隆起与横断山脉的纵横,把原本广泛分布的动植物种群分割开来,促成了许多独特的新物种诞生。又如海南岛与大陆间的琼州海峡,在历史时期的升降海面中反复隔离与连接,促进了岛屿与大陆生物的分化。
有些情况下,虽然两群体存在某种地理障碍,但依然有“边界地带”可以偶尔接触。这些地区常常出现杂交带,成为研究物种形成与基因流动的理想窗口。例如,秦岭地区不少动物的北方类型与南方类型就在杂交地带交流,这里的后代适应性往往较低。
同域物种形成
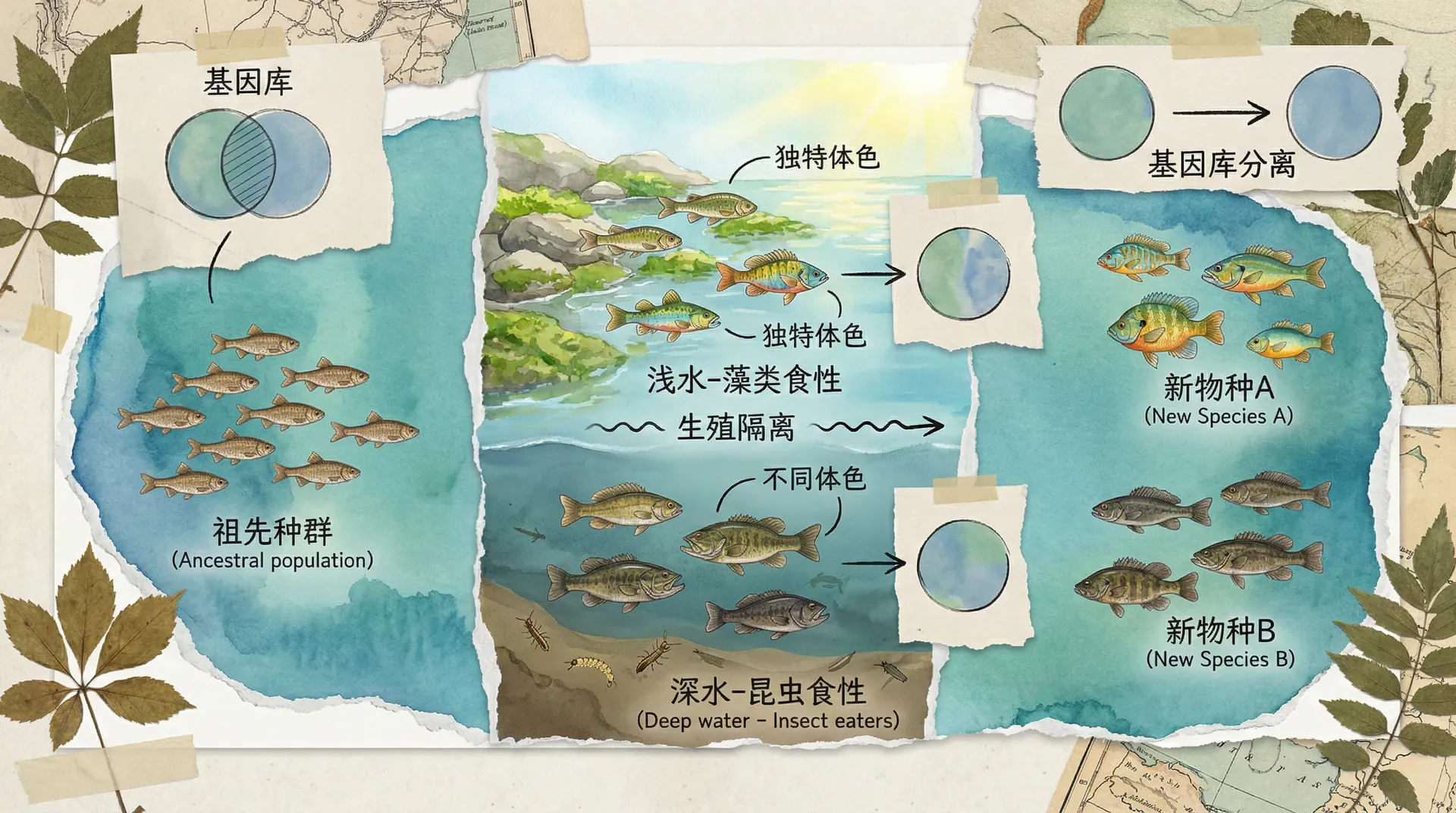
令人惊讶的是,物种分化并不总是依赖于地理隔离。许多情况下,在不发生空间分隔的前提下也能产生新物种,这被称为同域物种形成(sympatric speciation)。在植物中,尤其常见的机制就是染色体加倍——也称为“多倍体化”(polyploidization)。
当两个亲本物种杂交后,由于染色体配对的问题,初代杂种往往不能进行正常生殖,表现为不育。而一旦染色体加倍,新的配型又获得了生殖能力——这就是“异源多倍体”。这样产生的新物种通常兼具两个亲本的性状,并可在更加多样、复杂的新生态位中找到生长空间。中国的某些野生草本植物,如蕨类及部分禾本科植物,就有清晰的自然多倍体化物种形成的证据。
同域物种形成展现了生命进化的另一种可能性。它告诉我们,物种形成并不总是需要漫长的地理隔离过程,有时可以在生态压力和基因突变的共同作用下,在极短的时间和极小的空间范围内完成。这也让我们认识到物种界限本身具有一定的模糊性和流动性。
动物界的同域物种形成
虽然动物因配子形成、生殖行为等生理约束使得同域物种形成不如植物常见,但随着分子遗传学的发展,科学家们也陆续在部分昆虫、鱼类中发现了基于生态分化和行为隔离的同域物种形成案例。例如同一湖泊中不同食性的小型鱼种,可因选择不同资源最终形成繁殖隔离的新品系。
生殖隔离机制
无论是异域还是同域物种形成,进化的关键节点在于“生殖隔离”。只有当两个群体之间阻断了基因交流,才能真正成为彼此独立的物种。生殖隔离机制按照发生的时间可以分为合子前和合子后。合子前隔离防止受精发生,合子后隔离则表现为受精后杂种不能存活或繁殖。
下方总结了生殖隔离的主要类型及相关实例:
随着现代基因分析手段的发展,人们发现基因组水平的隔离比我们原先想象的还要复杂。例如,某些隔离性状只需区区一两个基因突变即可完成。
适应辐射
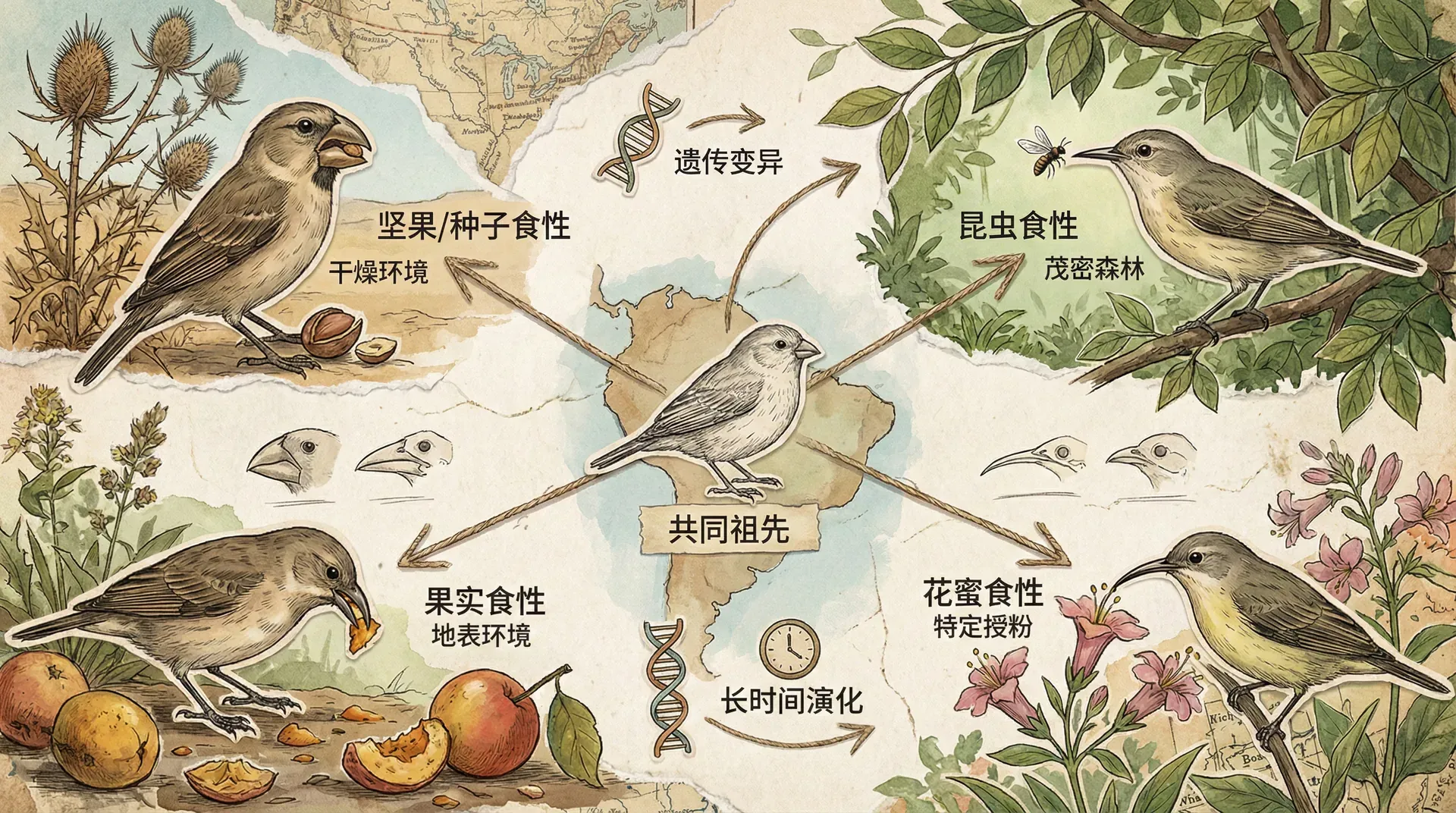
适应辐射(adaptive radiation)是指单一祖先在较短时间内演化出众多适应不同生态位的后代分支的现象。这一现象能迅速增加生物多样性,因此被认为是“生态机会”促成下的进化快车道。
青藏高原被誉为“世界屋脊”,其复杂的地貌和多样的微气候孕育了丰富的特有物种。从海拔较低的河谷到高耸入云的雪山,不同的环境条件塑造了差异明显的生物群落。这里不仅有高原特有的哺乳动物、鸟类,还有随着冰川变化反复扩散与隔离的高山植物。
垂直分布的物种格局反映出不同生物对温度、氧气、紫外线、植被类型等环境因素的适应能力。例如青藏高原的藏羚羊进化出高效的携氧能力与独特的体温调节机制,雪豹则拥有厚密皮毛和隐蔽的花纹,能够在极端气候中隐匿和狩猎。
世界各地的岛屿和高原都是适应辐射的天然实验田。例如加拉帕戈斯群岛的达尔文雀,不同岛屿上的雀类以种子、昆虫、花蜜为食,喙的大小形状沿着生态资源梯度出现了极快分化。我国云南、海南的岛屿型物种也展现了适应辐射的鲜活实例。
字符位移现象
当栖息于同一地区的相近物种相遇(称为“二重分布区”),往往产生形态或生态特征的序列性改变,以减少生态位重叠和种间竞争。这一演化过程被称为“字符位移”(character displacement)。
以中国南方山区的两种啄木鸟为例:当它们在各自单独分布时,喙的长度相差不大,资源利用高度重合。而当二者共存于山地混交林区时,其中一类的喙越发细长,主要取食树皮缝隙中的昆虫,另一类喙形更加粗壮,用以抓啄较深蛀虫。这种相互规避的生态适应,极大减少了物种间的竞争,促进了多样性的共存。
这种“字符位移”现象不仅见于啄木鸟,在许多鱼类、昆虫、青蛙、甚至鼠类等多类生物中反复出现。它表明生物群落的结构不是静态的,而是在种间相互作用、生态压力推动下不断动态演化。
字符位移现象提醒我们,竞争关系和生态环境的变化能够显著推动生物的分异与进化。这种共同进化(co-evolution)是自然界物种多样性和生态系统稳态的重要机制。
现代进化的观察
进化绝非只存在于远古时代,如今在我们身边的自然界甚至城市环境中,依然可以亲眼见证进化的发生。随着科技的发展,基因组学和长期的野外观测越来越多地揭示,“自然选择”正在日常生活中以肉眼可见或分子层面的方式持续进行着。
近年来,随着气候变化、城市化、土地开发和环境污染等人类活动不断加剧,动植物的生存环境发生了剧烈变化。在中国的城市生态研究中,科学家发现,一些鸟类为适应城市中不断增加的噪声环境,其鸣声的频率和节奏出现了明显变化,不同于郊区同类。这是城市适应进化的典型案例。类似地,诸如城市野鼠、杂草等动植物,也展现出了面对新环境的迅速遗传适应。农业中抗药性杂草、医院内抗生素耐药菌株的大量出现,更是现代进化的直观例证。生物通过新的基因突变或重组,快速获得了应对人类活动压力的生存策略。
理解进化的基本原理,不仅有助于我们深入认识生命多样性的本质,也让我们认识到人类在自然界中的真实位置。进化理论为解释分子、个体到生态系统各层级的生命现象,提供了统一的科学框架,也为保护生物多样性、应对全球环境变化奠定了坚实的基础。
进化是生命最根本、持久且充满创造力的特征。它连接着过去、现在和未来。通过研究进化的动力和过程,我们不仅能够解码地球生命的历史,还能更主动地预测和应对人类社会对自然环境的冲击,守护和促进全球生物多样性的持续繁荣。