种群生物学基础
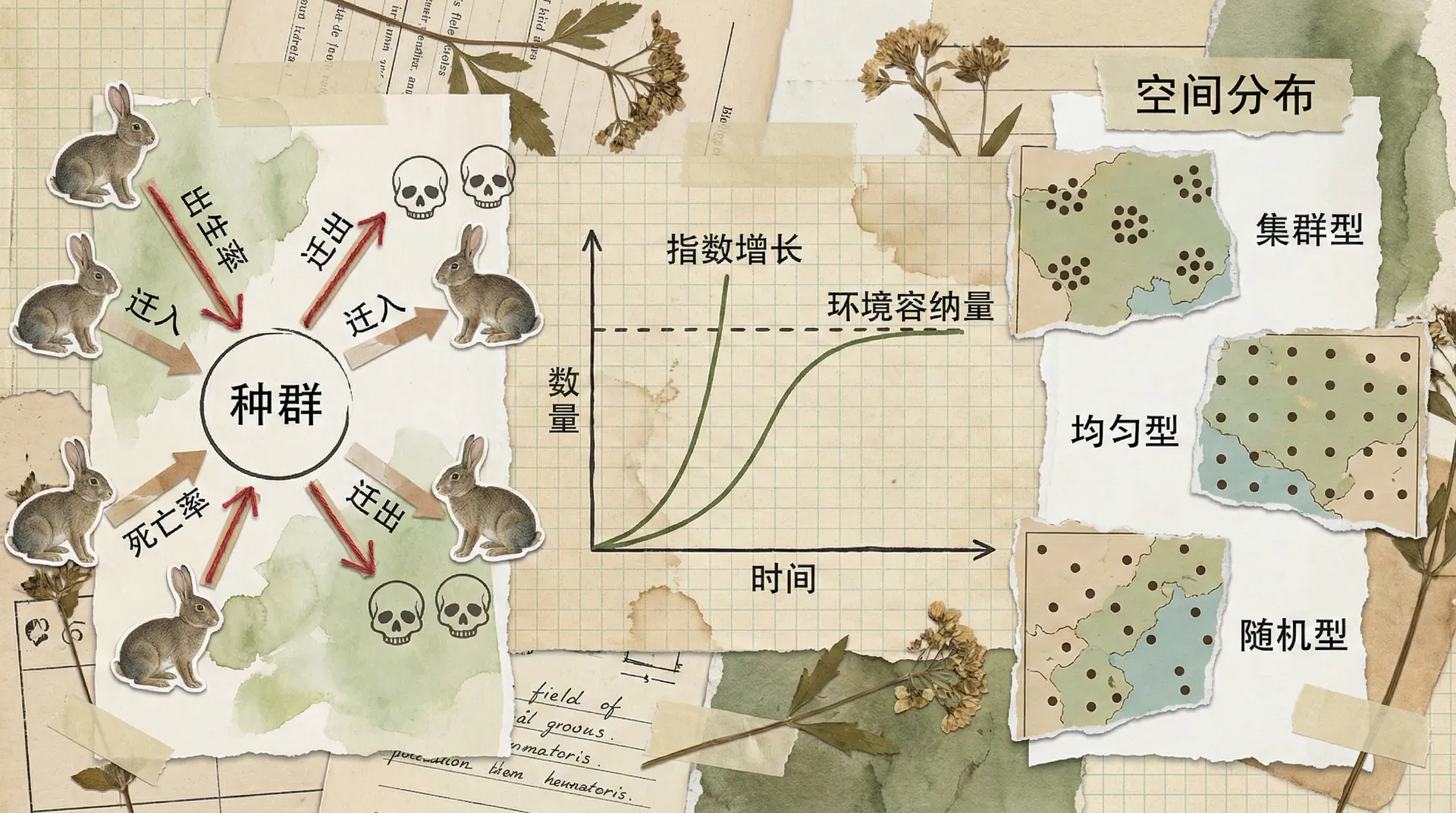
种群生物学(Population Biology)是研究生物种群数量变化及其规律的学科。它关注于同种生物在特定区域中所组成的种群,通过分析个体的出生、死亡、迁入与迁出等过程,揭示种群数量随时间和空间的动态变化。种群是生态系统的基本单位,不同物种的种群,其数量变化模式、生存策略和调节机制存在显著差异。
在自然界中,种群规模受到多种内外因素的共同影响,包括资源的供给、天敌的威胁、疾病流行和环境条件的变化。通过研究这些影响因素,种群生物学不仅能够解释种群数量的周期波动和突变,还能为野生动物保护、渔业管理以及入侵物种防控等实际问题提供理论基础和科学方法。
典型的问题包括:一个濒危动物种群能否恢复到安全水平?一个入侵种的扩散速度有多快?合理的资源利用强度应如何确定?这些都与种群的生存、繁殖、年龄结构、空间分布和与其他种群的相互作用密切相关。
现代种群生物学的发展与统计学、数学模型和分子生物学的结合日益紧密。通过生命表、种群增长模型和遗传分析等手段,科学家能够更加精细地刻画种群的历史与未来趋势。种群生物学的研究不仅丰富了我们对生物多样性维持机制的理解,也为生态文明建设和可持续资源管理提供了坚实的科学支撑。
生命表与种群统计分析
生命表是研究种群动态的重要工具,它系统地记录了不同年龄个体的生存和死亡情况。这个概念最初来自保险业,现在已成为生态学研究的核心方法。
生命表的基本原理
假如追踪1000只刚出生的丹顶鹤幼鸟的命运。第一年结束时,由于天敌捕食、疾病和环境压力,可能只有600只存活。第二年可能还剩400只,第三年300只,以此类推。这种记录就构成了生命表的基础数据。
生命表中的关键参数包括存活率、死亡率和生命期望。存活率lₓ表示从出生开始存活到x岁的个体比例。死亡率qₓ表示x岁个体在该年龄段内死亡的概率。生命期望Eₓ表示x岁个体平均还能存活的时间。
中国人口的生命统计特征
中国不同地区的人口生命统计数据展现了显著的地域差异。这种差异反映了经济发展水平、医疗条件和生活环境的影响。
这些数据说明了环境条件对种群统计参数的重要影响。发达地区由于医疗条件好、生活水平高,人口寿命更长,婴儿死亡率更低。
野生动物的生存分析
野生动物的生存分析为物种保护提供科学依据。以东北虎为例,通过无线电项圈跟踪和红外相机监测,科学家获得了珍贵的生存数据。例如,东北虎幼崽的第一年死亡率约为40%,主要死因包括天敌捕食、母虎死亡和栖息地破坏。成年东北虎的年死亡率约为10%,主要死因是人虎冲突、疾病和意外伤害。在良好保护条件下,东北虎的平均寿命可达15-18年。
通过生命表分析,保护工作者发现提高幼崽存活率是增加东北虎种群数量的关键。这导致了更多针对性的保护措施,如加强反偷猎巡护和改善栖息地质量。
生存的类型
不同物种表现出不同的生存模式,生物学家将其归纳为三种基本类型。
I型生存以东北虎为代表,特点是幼体和成体死亡率都较低,主要死亡集中在老年期。这种模式常见于大型哺乳动物和受到精心照料的物种。
II型生存以黄山猕猴为代表,各年龄段的死亡率相对恒定。这种模式在自然界比较常见,反映了稳定的生存压力。
III型生存以长江刀鲚等鱼类为代表,幼体死亡率极高,但一旦存活到成体阶段,死亡率就大大降低。这种策略通过大量产卵来补偿高幼体死亡率。
年龄结构的意义
种群的年龄结构反映了其发展趋势和健康状况。通过比较上海和西藏的人口年龄分布,我们可以看到不同发展阶段的人口特征。
上海的年龄结构呈现典型的老龄化特征,老年人口比例高,出生率低。西藏的年龄结构相对年轻,儿童和青年比例较高。这种差异不仅反映了经济社会发展水平,也预示着未来的人口发展趋势。
年龄结构失衡可能导致严重的社会和生态后果。对于野生动物保护来说,了解种群年龄结构有助于制定针对性的保护策略。
通过生命表分析和年龄结构研究,我们不仅能够了解种群的现状,还能预测其未来的发展趋势。这为科学管理和保护工作提供了重要的理论基础。
繁殖生物学与种群
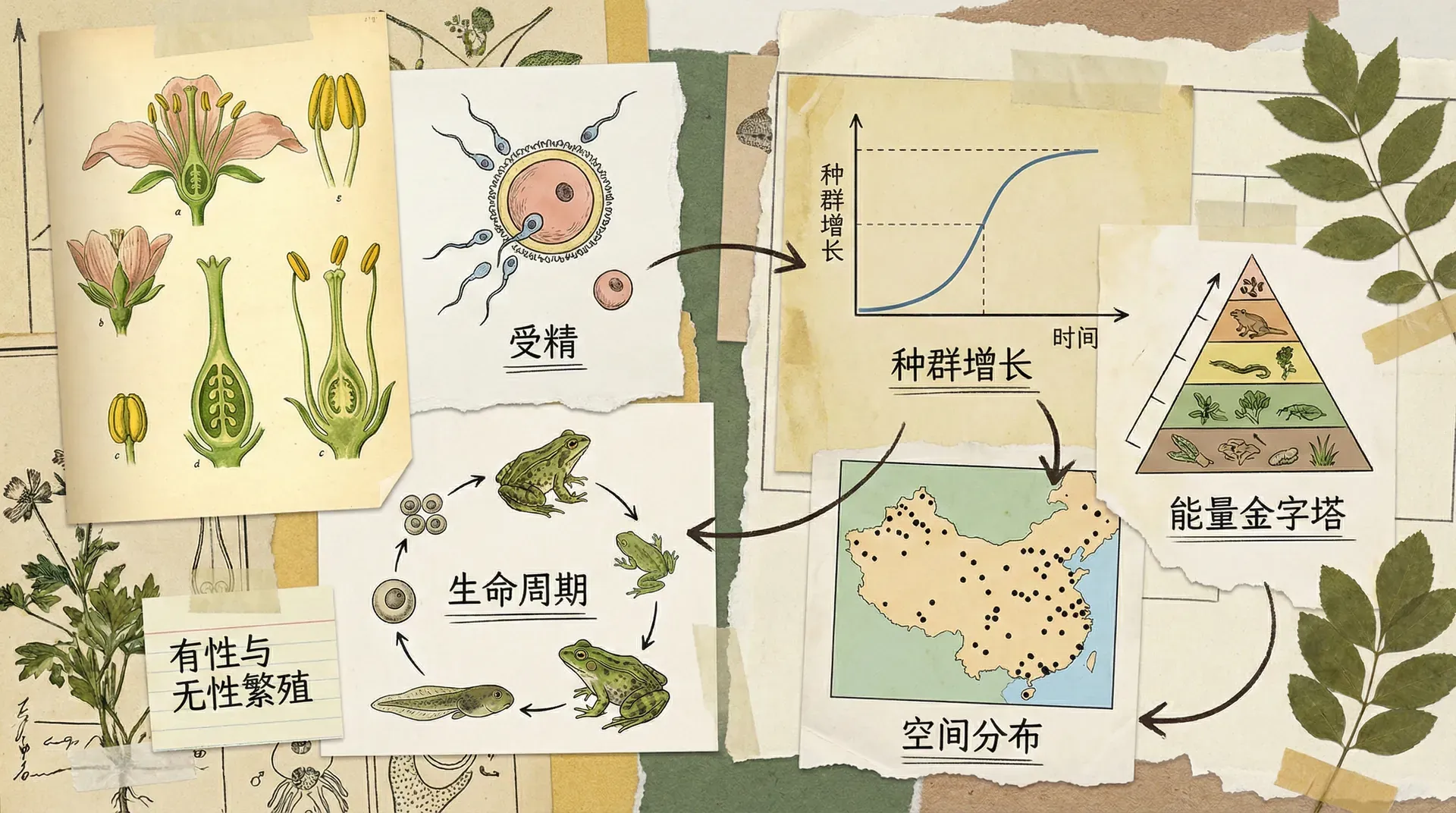
种群的持续存在依赖于成功的繁殖。不同物种采用了多样化的繁殖策略,这些策略在长期进化过程中得到优化,以适应特定的环境条件。
繁殖价值的概念
繁殖价值反映了不同年龄个体对种群未来发展的贡献潜力。以朱鹮为例,一只刚性成熟的3岁朱鹮具有很高的繁殖价值,因为它未来还有10多年的繁殖时间。而一只12岁的老龄朱鹮,虽然经验丰富,但繁殖价值相对较低。
繁殖价值是指某个年龄个体在其剩余生命中预期能够产生的后代数量,这个概念对制定保护策略具有重要指导意义。
幼鸟阶段繁殖价值为零,3-4岁达到性成熟后迅速上升,5-7岁达到峰值,之后随年龄增长逐渐下降。这种模式反映了生理状态和繁殖能力的变化规律。
净繁殖率与种群趋势
净繁殖率(R0)是衡量种群增长潜力的关键指标。它表示在当前的死亡率和繁殖率条件下,平均每个个体在一生中能产生多少个存活到繁殖年龄的后代。
当净繁殖率大于1时种群增长,等于1时种群稳定,小于1时种群衰减。这个简单的数字背后蕴含着种群命运的关键信息。以大熊猫为例,野生大熊猫的净繁殖率约为0.8-1.2,处在稳定和缓慢增长的边缘。人工圈养条件下,由于减少了天敌威胁和食物竞争,净繁殖率可以提高到1.5-2.0。
繁殖策略的多样性
自然界中的生物采用了截然不同的繁殖策略。毛竹代表了一次性繁殖(大爆发式繁殖)的极端例子,而银杏则是多次繁殖(迭代式繁殖)的典型代表。
毛竹的繁殖周期长达67年,在漫长的营养生长期后,整片竹林会在同一年开花结果,然后全部死亡。这种策略看似浪费,实际上具有重要的生态意义:同步开花可以避免种子被大量消耗,确保部分种子成功萌发。
相比之下,银杏采用多次繁殖策略,从50-100岁开始结实,可以持续几百年甚至上千年。每年的产种量相对较少,但总的繁殖投入分散在漫长的生命周期中,降低了单次繁殖失败的风险。
繁殖投入的权衡
生物在繁殖上的投入面临着复杂的权衡关系。过多的繁殖投入可能影响个体的存活,而投入不足则可能影响繁殖成功率。
朱鹮的人工繁育实践很好地说明了这种权衡。在人工饲养条件下,营养充足的朱鹮每年可以产3-4个蛋,但在野外自然条件下,通常只产1-2个蛋。更多的产蛋数量意味着需要更多的营养投入和更长的孵化时间,这在食物有限的自然环境中是有风险的。
这个对比表明,人工饲养条件下朱鹮可以承受更高的繁殖投入,但在自然环境中,适度的繁殖投入有利于雌鸟的长期存活。
后代质量与数量的平衡
繁殖策略不仅涉及投入多少资源,还涉及如何分配这些资源。是产生更多但较小的后代,还是产生较少但较大的后代?这是生物面临的基本选择。
藏羚羊的繁殖策略体现了对后代质量的重视。藏羚羊每胎通常只产一仔,妊娠期长达7个月,新生幼仔体重相对较大,约占母体重量的15%。这种策略确保了幼仔在严酷的高原环境中有足够的存活能力。相比之下,野猪采用了多仔策略,每胎产4-8仔,幼仔相对较小。这种策略通过数量优势来应对高幼仔死亡率的环境压力。
理解不同物种的繁殖策略差异,有助于制定更加精准的保护措施。对于藏羚羊这样的物种,保护重点应放在提高幼仔存活率上;而对于野猪等物种,则需要考虑栖息地容量的限制。
种群增长模式与调节机制

种群数量的变化遵循一定的规律。理解这些规律不仅有助于预测种群的未来趋势,也为资源管理和物种保护提供科学依据。
指数增长模型
在理想条件下,种群会表现出指数增长模式。这种增长的特点是:增长率恒定,但绝对增长量随时间加速增大。
水葫芦在中国的传播就是指数增长的典型例子。1901年作为观赏植物引入中国,最初几十年增长缓慢,但1980年代以后开始爆发式增长,现在已成为中国水域的主要入侵物种之一。
水葫芦的扩散过程清楚地展示了指数增长的三个阶段:缓慢建立期(1901-1980)、快速增长期(1980-2010)和趋于稳定期(2010至今)。
内在增长率的生物学意义
内在增长率(r)反映了种群在理想条件下的最大增长潜力。不同物种的r值差异巨大,这与它们的生活史特征密切相关。
这个表格显示了r值与生物特征的关系:体型小、成熟早、繁殖频率高的物种具有较高的r值,而大型的长寿物种r值较低。
环境阻力与种群调节
在自然条件下,种群的增长总会受到多方面的限制。以大熊猫为例,随着数量增加,受到栖息地面积、食物资源、个体间关系等因素的影响,增长速度逐渐减缓。最终,种群数量会稳定在一个由环境承载力决定的最大值附近,而不会无限制上升。
环境的承载上限决定了种群的最大数量。当种群逐渐接近这一上限时,常见的现象有:繁殖率降低、死亡率升高、种群成员发生迁移等。这种情况在很多受保护物种的管理中都值得关注。
种群波动与稳定性
即便接近环境极限,种群数量也不会一直保持静止,而是随着自然与外界条件起伏。例如,气候异常、食物短缺、疾病流行或人为干预等,都会引起数量的波动。
最近几十年,藏羚羊经历了数量快速下降、恢复增长到逐步趋于稳定的过程,这背后既有气候变化和天敌变化,也有保护措施的综合影响。稳定的生态系统,种群数量波动往往较小;而生态系统受扰动严重时,种群数量则可能出现较大起伏,甚至发生大幅降低。
掌握这些变化规律,有助于科学预测种群前景、优化物种保护和生态管理措施。在实际保护工作中,需要综合考量物种本身特性、环境周边条件及人为影响等各种因素。
衰老与死亡的进化意义
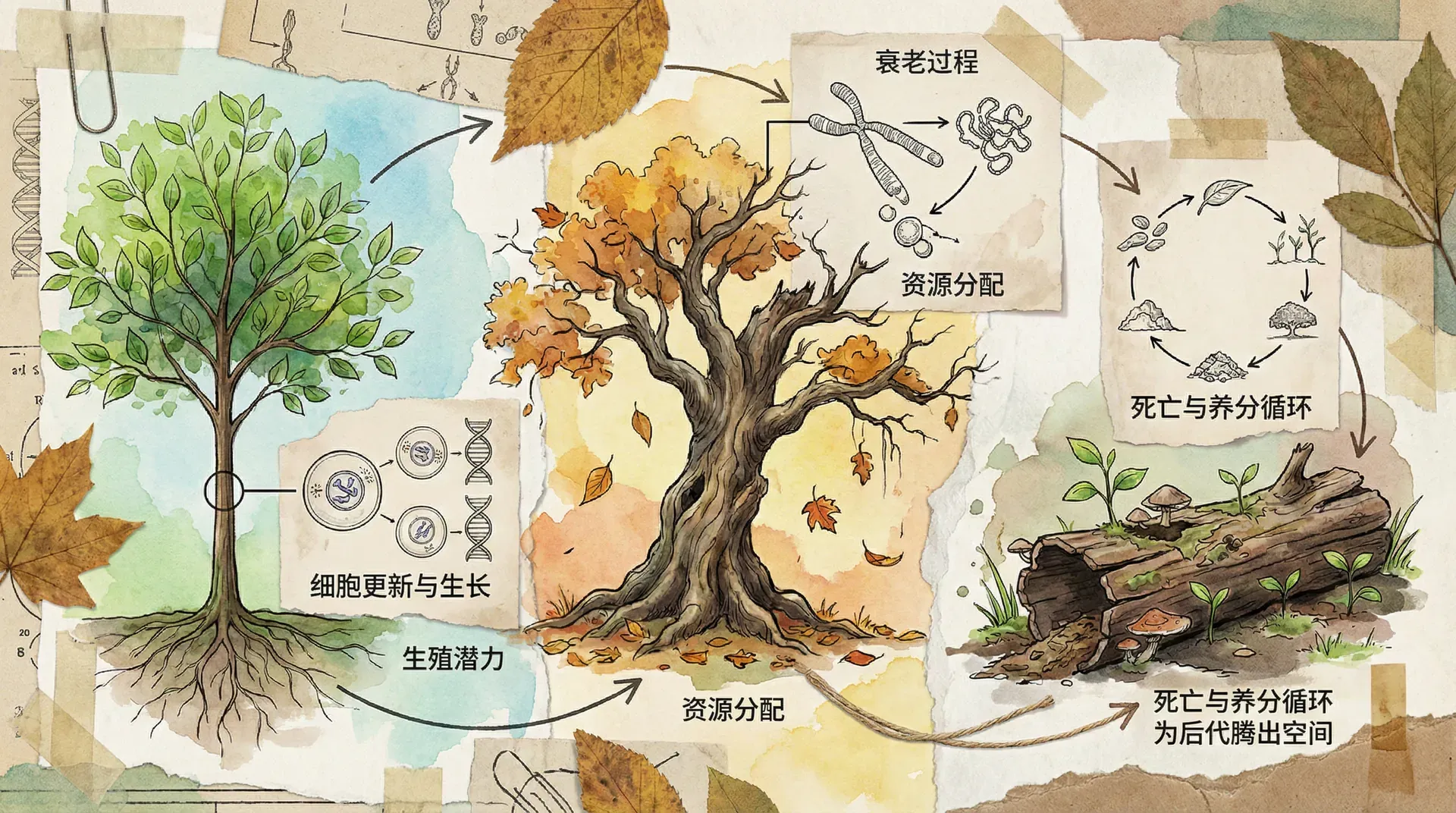
衰老是生物界的普遍现象,但为什么生物会衰老?从进化的角度看,衰老似乎是不利的,因为它降低了个体的生存和繁殖能力。然而,衰老现象在自然界中广泛存在,这说明它必然有其进化上的意义。
衰老的进化理论
现代进化生物学对衰老现象提出了几种解释理论。最被广泛接受的是繁殖价值递减理论:随着年龄增长,个体的繁殖价值逐渐降低,自然选择对晚期有害突变的选择压力也相应减弱。
黄山迎客松已有800多年历史,虽然现在仍然存活,但其生长势已明显衰弱,针叶稀疏,树冠不完整。从进化的角度看,这种衰老现象并非偶然。
在迎客松的前200年,它处于快速生长期,具有很高的繁殖潜力。自然选择强烈地筛选掉那些影响早期生长和繁殖的有害突变。但到了后期,即使出现影响老年健康的突变,由于此时的繁殖贡献已经很小,这些突变受到的选择压力也很弱,因此得以在种群中积累。
衰老不是进化的直接目标,而是自然选择在不同生命阶段作用强度差异的必然结果。这一认识对理解长寿物种的保护策略具有重要意义。
中国古树群落的衰老模式
中国拥有丰富的古树资源,这些古树为研究植物衰老提供了珍贵材料。不同物种的古树展现出不同的衰老模式和适应策略。
银杏是著名的“活化石”植物,单株寿命可达数千年。山东莒县的银杏古树群中,树龄超过1000年的银杏仍能正常开花结果,这说明银杏具有独特的抗衰老机制。
相比之下,胡杨虽然也是长寿树种,但其衰老模式与银杏不同。塔里木河流域的千年胡杨林显示,胡杨通过不断萌发新枝来延续生命,老枝死亡而新枝继续生长,形成了特殊的“更新式衰老”模式。
动物的衰老与寿命进化
动物的衰老模式比植物更加复杂多样。在中国的野生动物中,不同物种的寿命和衰老特征差异巨大,这些差异反映了不同的进化策略。
东北虎的野外寿命约为15-18年,人工饲养条件下可达20-25年。东北虎的衰老主要表现为狩猎能力下降、牙齿磨损、肌肉量减少。这种衰老模式与其作为顶级掠食者的生活方式密切相关:一旦狩猎能力严重下降,个体就难以在野外生存。
相比之下,大熊猫的寿命相对较长,野外约20年,人工饲养可达35年以上。大熊猫的衰老过程相对缓慢,主要表现为消化能力下降、活动减少、毛色变化。这种缓慢衰老可能与其相对简单的生活方式和低代谢率有关。
这个对比显示了环境条件对动物衰老和寿命的重要影响。圈养条件下,消除了天敌威胁、食物短缺等生存压力,动物的寿命可以显著延长。
衰老速度的种间差异
不同物种的衰老速度差异巨大,这些差异与其生活史策略密切相关。一般而言,体型大、代谢率低、繁殖开始晚的物种寿命更长,衰老速度更慢。
在中国的哺乳动物中,这种模式表现得很明显:
这种体重-寿命关系反映了代谢理论的预测:大型动物具有较低的质量特异性代谢率,细胞分裂频率较低,DNA损伤积累较慢,因此寿命更长。
环境压力与衰老适应
环境压力显著影响生物的衰老进程和寿命。在严酷环境中生存的物种往往演化出了特殊的抗衰老机制。
藏羚羊生活在海拔4000米以上的青藏高原,面临缺氧、低温、强紫外线等多重环境胁迫。在这种环境中,藏羚羊演化出了强大的抗氧化系统和DNA修复机制,使其能够在极端环境中维持相对较长的寿命(野外约12-15年)。
塔里木胡杨林中的胡杨面临极度干旱的环境,年降水量不足100毫米。在这种条件下,胡杨通过降低代谢率、增强细胞修复能力等方式来延缓衰老,单株寿命可达千年以上。
极端环境中的物种往往具有独特的抗衰老机制,这些机制不仅帮助它们在恶劣环境中生存,也为人类抗衰老研究提供了重要启示。
衰老与种群保护
理解衰老的进化机制对野生动物保护具有重要意义。在制定保护策略时,需要考虑不同年龄个体对种群贡献的差异。
对于大熊猫这样的长寿物种,老龄个体虽然繁殖能力下降,但它们的生存经验对种群仍有价值。老龄雌性大熊猫往往是优质栖息地的占据者,它们的行为模式会影响年轻个体的栖息地选择和生存策略。相反,对于东北虎这样的短寿种,繁殖期个体的保护优先级更高。一只处于繁殖高峰期的雌虎对种群的贡献远大于一只老龄雌虎,因此在资源有限的情况下,应优先保护繁殖期个体。
基于衰老生物学的保护策略需要平衡不同年龄个体的生态价值和繁殖贡献,这种精细化的保护理念正逐渐被应用于珍稀物种的保护实践中。
总结
种群生物学不仅为物种保护、资源管理和生物入侵防控等领域提供了坚实的科学理论基础和有效的分析工具,还推动了相关实际工作的不断优化。例如,中国朱鹮的濒危物种保护与恢复,就是在深入研究种群动态、遗传多样性及其栖息地微环境变化的基础上,通过精细化管理和科学干预,最终实现了朱鹮种群从边缘灭绝到健康增长的典型转变。
在资源管理方面,野生动物种群的数量监测、年龄结构分析及繁殖率评估等工作,为可持续利用和合理开发提供了重要决策依据。对入侵物种而言,通过构建其扩散模型和评估种群参数,可以制订出更具针对性的防控和治理策略。随着大数据、遥感、分子生物学等交叉学科方法的不断应用和创新,种群生物学将在未来的生态文明建设、环境管理及全球生物多样性保护中展现出更为广阔的应用前景和影响力。