生态竞争

生态系统中的生物并不是孤立存在的,而是彼此相互联系、相互影响的整体。它们在生命活动过程中持续发生着多种多样的相互作用,其中竞争被认为是最为重要且普遍存在的一种关系。竞争不仅发生在野外的原始森林和草原中,也广泛出现在我们身边的各种自然和人工生态系统里。
比如,在城市公园中,各类植物为了争夺阳光、水分和养分而展开激烈的竞争;在农田里,作物和杂草之间也在无时无刻不进行着生存资源的分配和争夺。即使是在微观世界中,如土壤中的微生物,也因空间或营养来源的限制进行着看不见的激烈竞争。正是这种无声却持久的生存较量,推动着生物种群的动态变化,并影响着生态系统的结构与稳定性。
什么是生态竞争
竞争的基本概念
生态竞争是指两个或多个生物个体或群体为了获取有限的资源而产生的相互作用过程。这些资源可能是食物、栖息地、阳光、水分,或者配偶。当资源充足时,生物之间基本不存在竞争;但当资源变得稀缺时,竞争就会变得激烈起来。
竞争并不是一种“非有即无”的现象,而是一个连续变化的过程。竞争的激烈程度取决于资源需求量与供给量的比值。
我们可以把竞争分为两大类型:种内竞争和种间竞争。种内竞争发生在同一物种的个体之间,比如同一片森林中的松树为阳光而竞争;种间竞争则发生在不同物种之间,比如水稻和稗草争夺土壤中的养分。
竞争的三种方式
生物进行竞争的方式可以分为三种基本类型:
消耗性竞争是最常见的竞争方式,也称为利用性竞争。在这种竞争中,生物通过更快或更有效地消耗共同需要的资源来获得优势。比如在中国北方的草原上,不同种类的草本植物都需要土壤中的氮、磷等营养元素,谁能更快速地吸收这些养分,谁就能获得更好的生长条件。
干扰性竞争是一种更加直接的竞争方式,涉及生物之间的直接对抗。这可能表现为物理攻击、化学抑制或者领域行为。例如,胡杨树会分泌化学物质抑制其他植物在其附近生长,这种现象在我国西北地区的荒漠绿洲中很常见。
抢占性竞争主要围绕空间资源进行,谁先占据了有利位置,谁就获得了竞争优势。在潮间带的岩石表面,各种海藻和贝类为了附着空间展开激烈竞争,这在我国东南沿海地区的海岸生态系统中表现得尤为明显。
上图显示了竞争强度与资源可获得性之间的关系。可以看出,当资源充足时,生物之间几乎不存在竞争;随着资源变得稀缺,竞争强度急剧上升。
种内竞争与种间竞争

种内竞争的特点
种内竞争是同种生物个体之间的竞争,这种竞争往往最为激烈,因为同种个体对资源的需求几乎完全相同。在河南省的小麦田中,我们可以观察到明显的种内竞争现象:当小麦播种密度过高时,幼苗之间为了获取阳光和养分而竞争,结果导致植株变细、产量下降。
种内竞争有一个重要特点,就是它具有自我调节的功能。当种群密度过高时,激烈的竞争会导致生存率下降,从而使种群密度回到合适的水平。这种现象在许多动物种群中都能观察到,比如东北地区的野猪种群。
种间竞争的作用
种间竞争的结果往往更加复杂多样。在长期的进化过程中,种间竞争成为推动物种分化和生态位分离的重要力量。当两个物种的生态需求完全重叠时,通常会出现竞争排斥现象,即其中一个物种会被另一个物种完全排挤出去。
但是,种间竞争也可能促进物种的共存。通过长期的相互适应,不同物种可能在时间、空间或者食物选择上发生分化,从而减少直接竞争,实现和谐共存。
竞争强度的影响因素
竞争强度受到多个因素的影响,其中最重要的是环境容量和种群密度。在四川省的竹林生态系统中,我们可以观察到这样的现象:在养分丰富的山谷底部,不同种类的竹子可以和谐共存;但在贫瘠的山坡上,竞争变得更加激烈,往往只有少数几种适应性强的竹种能够存活。
竞争排斥现象
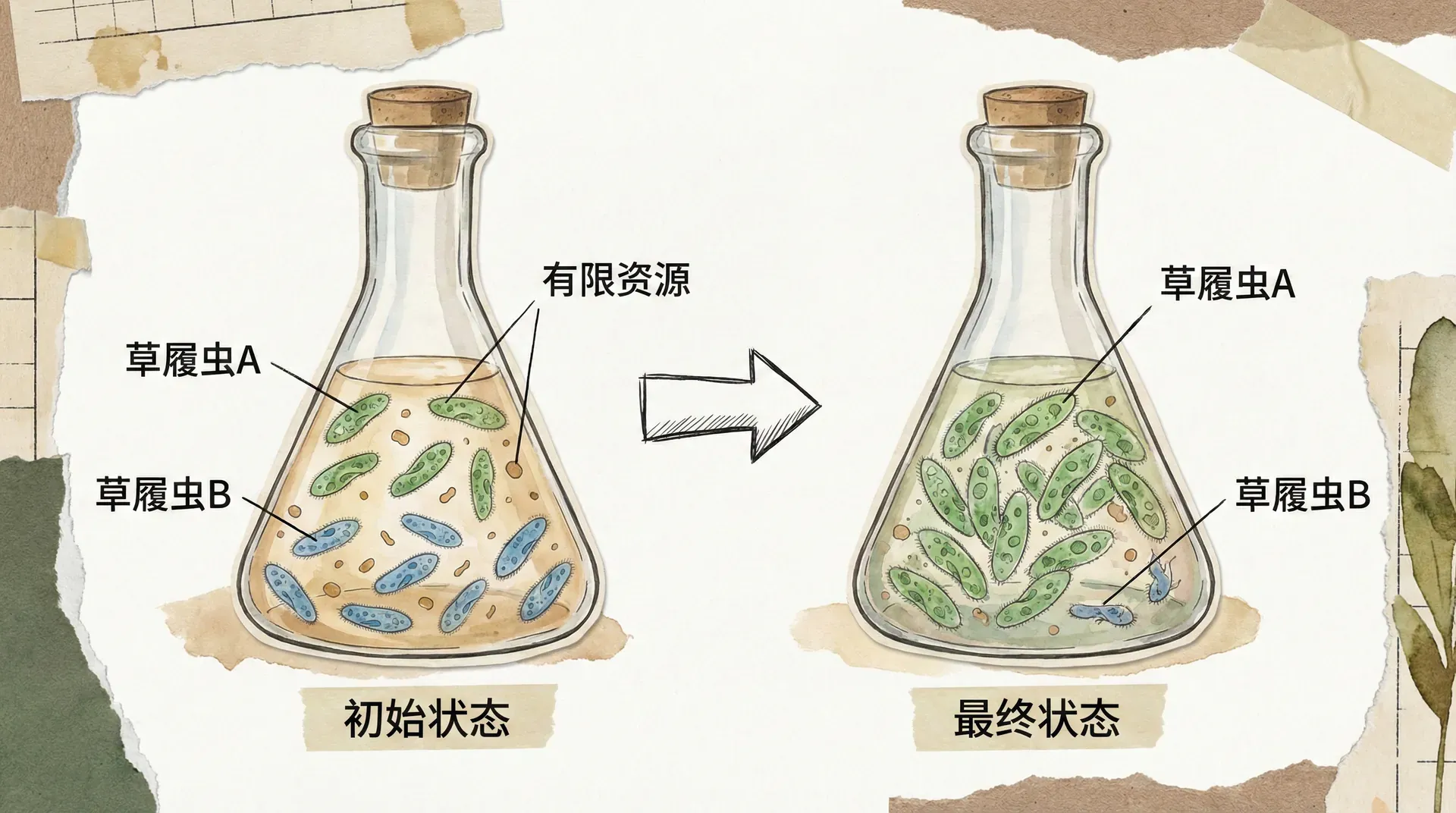
竞争排斥的过程
竞争排斥是生态学中一个重要概念,描述的是当两个生态需求完全相同的物种同时存在于同一环境中时,其中一个物种会被另一个物种完全排除的现象。这个过程通常需要经历几个阶段。
最初,两个物种都能够在环境中正常生长繁殖。但随着种群数量的增加和资源的日益稀缺,竞争开始加剧。由于两个物种不可能在所有方面都完全相同,必然会有一个在某些方面更有优势。随着时间推移,这种微小的优势会被不断放大,最终导致劣势物种的完全消失。
中国的入侵物种
加拿大一枝黄花的入侵就是竞争排斥现象的例子。这种原产北美的植物在20世纪30年代被引入我国作为观赏植物,但很快就逃逸到野外。加拿大一枝黄花具有极强的繁殖能力和竞争优势,它能够快速占领空地,并通过地下根茎迅速扩张。
加拿大一枝黄花之所以能够成功入侵,主要因为它在新环境中缺少天敌,同时具有比本土植物更强的资源获取能力和更快的生长速度。
在江浙一带的农田边缘和荒地中,加拿大一枝黄花大面积生长,排挤了许多本土植物,如野菊、紫云英等。这种单一物种的大面积扩张不仅破坏了植物群落的多样性,还影响了以这些本土植物为食的昆虫和其他小动物的生存。
水葫芦(凤眼莲)是另一个典型的例子。这种原产南美的水生植物在我国南方的湖泊和河流中大量繁殖,形成了厚厚的绿毯,阻断了阳光对水下植物的照射,同时消耗了水中的氧气,导致鱼类和其他水生动物死亡。
竞争结果的四种可能性
生态学家通过理论分析和实验观察,总结出种间竞争可能产生的四种结果:
稳定共存是最理想的结果,这要求每个物种对自身种群增长的抑制作用要强于对其他物种的抑制作用。换句话说,种内竞争必须强于种间竞争。
| 初始物种A密度 | 初始物种B密度 | 竞争结果 |
|---|---|---|
| 高 | 低 | A物种获胜 |
| 低 | 高 | B物种获胜 |
| 中等 | 中等 | 共存区域 |
| 接近边界 | 接近边界 | 结果依赖初始分布,存在不稳定共存 |
不同的初始密度条件下,两物种的竞争结果可表现为A获胜、B获胜、稳定共存,或依赖初始情况的不稳定共存。共存通常出现在两物种对自身的抑制作用大于对方时。
竞争的平衡机制
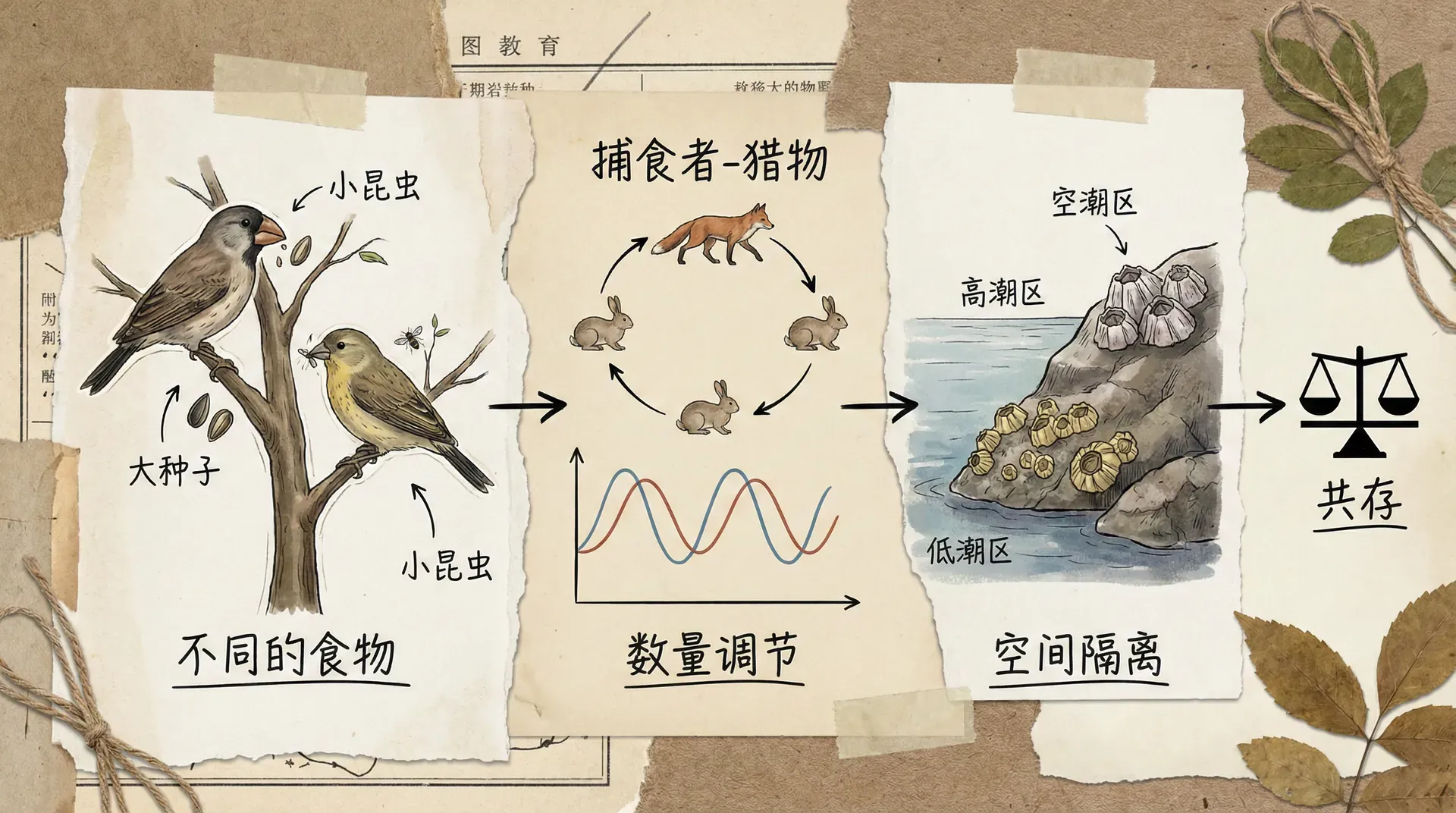
栖息地选择与资源分配
在自然界中,生物很少会盲目地争夺相同的资源,而是会根据环境条件和竞争压力来调整自己的行为。栖息地选择就是一种重要的竞争缓解机制。
以东北地区的森林为例,在同一片林地中生长着多种鸟类,它们虽然都以昆虫为食,但在空间利用上却表现出明显的分化。啄木鸟主要在树干上觅食,山雀类更偏好在树枝间活动,而地面的鸟类如鸫则主要在林下层寻找食物。
这种空间分化的形成遵循一个重要原理:当所有个体都选择最优栖息地时,由于竞争的加剧,这些"最优"栖息地的实际价值会下降;相反,一些看似次优的栖息地由于竞争较少,反而能为个体提供更好的生存条件。
生态位分化
生态位分化是指不同物种在长期的共同进化过程中,逐渐在资源利用、栖息地选择或活动时间等方面产生差异,从而减少直接竞争的现象。
在中国的暖温带落叶阔叶林中,我们可以观察到树种间明显的生态位分化。例如,杨树通常占据阳光充足的开阔地带,而槭树则更适应林缘的半阴环境,橡树能够忍受更为阴蔽的林下条件。这种垂直空间的分化使得不同树种能够在同一森林中和谐共存。
鸟类的取食生态位分化更是精细入微。在长江中下游地区的湿地中,不同种类的鸟类虽然都以鱼类为食,但它们的喙长、腿长和取食行为都有所不同。白鹭具有长喙和长腿,适合在深水中捕鱼;苍鹭更加高大,能够涉入更深的水域;而白鹮的弯曲长嘴则特别适合在泥滩中探寻小鱼虾。
时间和空间的分离
时间分离是另一种重要的竞争缓解机制。许多生态需求相似的物种通过在不同时间进行活动来避免直接竞争。
昼夜活动节律的差异是最常见的时间分离形式。在中国北方的农田生态系统中,麻雀和蝙蝠虽然都以昆虫为食,但麻雀在白天活动,而蝙蝠在夜间出没,从而避免了食物竞争。
季节性的活动差异也很重要。在青藏高原的高山草甸中,不同植物的开花时间错开分布:早春时节雪莲率先开花,夏季中期是龙胆花的盛花期,而秋季则是紫菀类植物的开花季节。这种时间上的分离减少了对传粉昆虫的竞争。
竞争塑造的进化适应
性状替代现象
性状替代是指两个相近的物种在同域分布时比在异域分布时表现出更大差异的现象。这种现象被认为是竞争压力推动进化的直接证据。
在中国东部沿海地区的潮间带生态系统中,我们可以观察到贝类的性状替代现象。研究发现,两种相近的牡蛎在各自单独分布的区域内,它们的壳型和大小都比较相似;但在两者共同分布的区域,这两种牡蛎的形态差异显著增大,一种趋向于扁平宽阔的壳型,另一种则发展出更加厚实紧凑的壳型。
鸟类的喙部形态也展现出明显的性状替代。在海南岛的热带雨林中,当两种相似的啄木鸟同时存在时,它们的喙长差异比在各自单独分布区域时更加明显。这种差异使得它们能够开发不同深度的虫洞,从而减少食物竞争。
形态差异与生存策略
长期的竞争压力不仅塑造了物种的形态特征,还影响了它们的行为策略和生理特征。
在中国西南地区的喀斯特地貌中,不同种类的攀援植物发展出了各具特色的攀爬策略。木质藤本植物如猕猴桃依靠强健的茎干缠绕上升,而草质藤本如葛藤则通过快速生长和密集分枝来抢占光照空间。一些植物如薜荔发展出了气生根,能够在垂直岩壁上攀爬,开辟了其他植物无法利用的生态空间。
上图显示了两个相近物种在同域分布时比异域分布时表现出更大的形态差异,这是竞争驱动进化的重要证据。
中国物种的进化
中国幅员辽阔,地形复杂,为物种的适应性进化提供了丰富的舞台。大熊猫和小熊猫就是一个有趣的例子,虽然它们都以竹子为主食,但在食物选择上表现出了明显的分化:大熊猫主要取食竹子的嫩茎,而小熊猫更偏好竹叶,这种取食差异减少了两者之间的食物竞争。
在青藏高原上,不同海拔高度的植物群落展现出了明显的垂直带状分布。随着海拔升高,温度下降,氧气稀薄,植物逐渐从高大的乔木过渡到低矮的灌木,最后是贴地生长的草本植物。每一个海拔带的植物都发展出了适应当地环境的特殊形态和生理特征。
竞争不仅是一种破坏性力量,更是推动生物多样性发展的重要动力。正是在竞争的压力下,不同物种才能够不断进化出新的适应策略,最终形成丰富多彩的生物世界。
竞争研究的科学证据
实验室观察结果
科学家在实验室条件下进行的竞争实验为我们理解竞争机制提供了重要证据。中国科学院的研究人员曾经用两种常见的水蚤进行了经典的竞争实验。
实验设计很简单:在相同的培养条件下,分别单独培养两种水蚤,然后将它们放在一起混合培养,观察种群数量的变化。结果发现,当单独培养时,两种水蚤都能稳定增长并维持在较高的数量水平;但当混合培养时,其中一种水蚤的数量逐渐下降,最终完全消失。
华南农业大学的研究团队还进行了不同环境条件对竞争结果影响的实验。他们发现,在温度和湿度不同的条件下,同样两个物种的竞争结果可能完全不同。这说明环境因子对竞争强度和竞争结果有重要影响。
野外调查发现
野外的竞争现象往往更加复杂,但也为我们提供了更接近真实情况的观察结果。
在长白山的针阔混交林中,科研人员发现了鸟类觅食行为的精细分化。五种外形相似的鸟类虽然都在同一片森林中生活,都以昆虫为食,但它们在树冠的不同部位进行觅食活动。有的主要在树冠顶部活动,有的偏好树干中部,还有的专门在枝条末梢寻找食物。
这种空间分化如此精确,以至于研究人员能够根据鸟类在树冠中的位置来预测它们的种类。更有趣的是,当其中一个物种因为迁徙或其他原因暂时离开时,其他物种会扩大自己的活动范围,临时占用空出来的生态空间。
在中国南方的水稻田生态系统中,研究人员发现了植物与昆虫之间复杂的竞争关系网络。水稻不仅要与杂草竞争阳光、养分和空间,还要面对多种害虫的取食压力。而这些害虫之间也存在复杂的竞争关系,比如稻飞虱和稻蓟马虽然都取食水稻汁液,但它们偏好植株的不同部位,从而减少了直接竞争。
中国生态保护的应用
竞争理论在中国的生态保护和恢复工作中发挥着重要作用。
在退耕还林工程中,科学家利用竞争原理来设计植物群落的搭配。例如,在西北地区的荒山造林中,会同时种植不同根系深度的树种:深根系的刺槐能够利用深层土壤水分,而浅根系的沙棘则主要利用表层水分,这样可以提高水资源的利用效率,减少种间竞争。
在生物防治领域,科学家利用天敌昆虫与害虫之间的竞争关系来控制农业害虫。比如,通过释放寄生蜂来控制玉米螟的危害。寄生蜂的幼虫在玉米螟体内发育,最终导致玉米螟死亡,这种生物防治方法既环保又有效。
在生态恢复工作中,必须充分考虑物种间的竞争关系。盲目引入外来物种可能会破坏本土生态系统的平衡,造成不可逆转的生态损失。
生态竞争的意义
理解生态竞争机制,有助于保护生物多样性和提升生态管理水平。许多濒危物种面临的竞争压力,特别是来自外来物种的入侵,已成为生物多样性保护的重点问题。例如,中华鲟与其他鱼类竞争有限的食物资源,大熊猫要依赖于竹子的持续更新,保护工作都需统筹群落平衡,防止某类资源被过度占据。
在农业领域,利用作物和天敌之间的互补和竞争关系,可以减少农药使用,提高产出,如玉米与大豆套种、生物防治棉蚜等做法。覆盖作物通过抢占资源抑制杂草,同时改善土壤,充分体现了竞争机制的积极作用。
城市生态建设也需充分考虑植物的竞争关系。合理搭配乔木、灌木、草本,分层配置植被,有效利用空间并提升生态效益,促进稳定的城市绿色群落。例如,不同植物适应不同水深和光照,通过科学配置支持整体生态系统的健康。
深入理解生态竞争,有助于实现资源的合理配置和环境的可持续发展。只有科学规划和协调,才能促进自然与社会的共同繁荣。每个物种都在生态系统中有其独特作用,保护生物多样性,就是守护我们的未来。