生态位
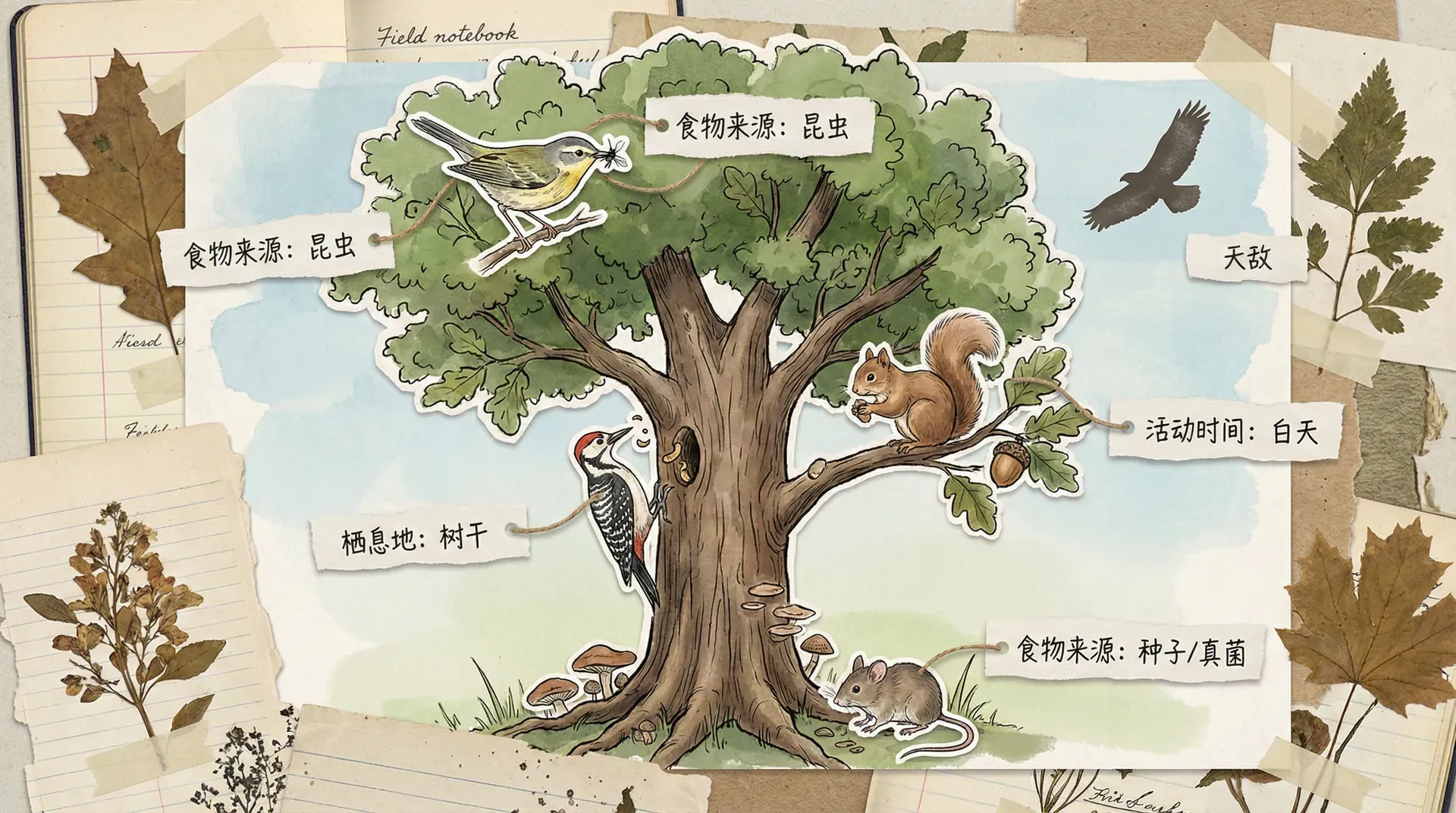
在大自然这个庞大而复杂的“社会”中,每种生物都像拥有独特身份证明的成员一样,拥有各自专属的生存方式和生活空间。就像人类社会中每个人有自己的职业和家庭住址,不同的生物也在生态系统中扮演着不同的角色,适应着属于它们自己的“位置”。生物学家将这种现象称为“生态位”。所谓生态位,实际上是在描述生物如何在纷繁多样的自然环境中,找到自己独特的位置——既包括它们选择什么样的“家”,又包括它们如何与周围的环境和其他生物发生联系。
生态位概念的发展历程
生态位(Niche)不仅是生物在生态系统中占据的位置,更强调它们在系统中扮演的功能角色和生存地位,包括对环境资源的利用方式、与其他生物的互作关系、以及对自然环境变化的响应策略。生态位的概念贯穿了生物的“吃、住、行、社交”等方方面面。
在生态学的发展历程中,科学家对“生态位”有着多元而丰富的解读。早期的生物学家们各执一词,就像“盲人摸象”般从不同角度理解和诠释这个复杂但又极具包容性的概念。随着研究的深入,生态位理论不断充实和完善,逐渐成为生态学研究的核心理论之一。我们可以通过下面的表格对比几位关键学者关于生态位的经典观点:
其中,奥德姆的比喻最为生动。他将“栖息地”比作生物的“家庭住址”,而“生态位”则等同于它们的“职业特长”。举个具体的例子,比如大熊猫:它的住址是中国四川的茂密竹林,而它的“职业”则可以说是“竹子专业清理工”——不仅因为它以竹子为主食,还因它对竹林生态系统的维持具有独特作用。
当然,不同生物可能在生态系统中的“职业”会发生变化,甚至同一种生物在不同环境或不同阶段也会调整自己的“角色”。比如部分鸟类可能在繁殖季节时拥有领地行为,而在非繁殖季则变得群居。这种灵活的生态位占据方式也是生物多样性维持与生态系统复杂性的重要基础。
哈钦森的超维度生态位模型
要全面描述一个人的生活状态,只关注单一维度远远不够,我们往往需要综合考虑收入水平、居住环境、工作地点、兴趣爱好、作息规律、家庭背景、社交圈子等诸多因素。每一个维度都可能影响这个人在社会中的“定位”以及能参与的各种活动。类似地,要全面刻画一个物种的生态位,也必须将温度、湿度、光照、食物类型、栖息高度、天敌压力、繁殖时间、竞争对手、资源获取方式等各种生态因子纳入考量。正是这些变量的综合作用,共同界定了物种能够生活的“生态空间”——这正是哈钦森提出的“超维度生态位”概念的核心。
在这个多维空间的示意图中,蓝色椭圆代表“基础生态位”(Fundamental Niche):假如完全没有外来干扰(比如天敌、竞争者、疾病或人为破坏等),物种理论上能够存活和繁衍的环境范围;而红色椭圆则是“实现生态位”(Realized Niche):在现实生态系统中,受种间竞争、捕食关系、资源限制等实际生态压力影响后,物种真正能够成功立足的空间通常变得更窄、更有限。
基础生态位与实现生态位
基础生态位指的是物种在最理想、无其它生物干扰的条件下,能够生活的所有可能环境范围。
实现生态位是实际生态系统中,物种在竞争、捕食和其他生态因子影响下,实际被“挤压”出来的、可以维持其种群的环境区域。
这种差异就如同一个拥有多项技能和学历的大学毕业生,按理说可以胜任许多种职业(基础生态位),但由于现实社会的职位竞争、用人单位的需求筛选等客观限制,最终他能实际获得并长期从事的职业(实现生态位)要少得多,受到了种种约束。
具体到自然界,例如,中华大蟾蜍理论上可以适应多种湿润环境、分布范围很广(这就是它的基础生态位),但实际上,由于受到其他蛙类间的竞争,以及湿地环境的破坏和缩小,它们能够真正占据的生境往往被限制在一些特定的池塘和湿地中(对应实现生态位)。同样道理,许多湖泊鱼类能够忍耐较宽的温度和盐度波动,但实际生态系统里受到捕食压力或食物资源的限制后,就只能在有限的水体区域生存。
哈钦森的生态位模型把生物的“生存空间”拓展到了多维环境变量的抽象空间,让我们能够系统地比较、量化不同生物之间的生态位差异与重叠,这对于理解物种多样性形成、物种间竞争关系乃至于生物多样性保护都有重要意义。
生态位重叠与种间竞争

当两个物种需要相同或高度相似的资源时,就会发生生态位重叠。这在自然界中极为普遍——例如不同鸟类利用大小相近的食物、鱼类在水体不同深度中觅食、以及森林里多种树种争夺阳光和土壤养分。生态位重叠并不仅仅体现在食物上,还可以在空间(栖息地)、时间(活动时间)、筑巢地点等多个维度发生。由于生态系统中的资源往往有限,生态位重叠通常是激烈竞争的起点。
从上图可以看出,喜鹊和乌鸦在4-6毫米大小的食物资源利用上存在重叠。也就是说,这两个物种在这部分区间内会“抢食”,此时谁的捕食效率高、耐饥饿更强或行为更敏捷,谁就更可能获得成功。此外,在生态位边缘部分,两个物种各有专长,可以有效利用不同大小的食物资源,从而部分减小竞争压力。
生态位重叠的现实例子还有很多,如中国北方的褐家鼠和小家鼠均会在人类建筑物中活动,争夺同样的食物和空间资源;河流或湖泊中的鲤鱼和鲫鱼也会争夺底栖无脊椎动物作为食物。
竞争的结果模式
生态位重叠导致的种间竞争,可能出现多种结果:
生态学上把不同物种能够共存的机制总结为“生态位分化”或“资源分配”—也就是各自“错开”生态位,减少重叠。例如许多鸟类会分不同时间觅食、或专注于特定的食物类型,从而避开激烈的竞食。还有的物种在长期进化中改变生理结构、行为方式,逐步缩小重叠范围,最终和平共处。
重要的是,生态位重叠仅在资源稀缺时才诱发激烈竞争,若资源充足,则重叠的物种可以共存。但若资源极端匮乏,那么弱势者往往会被排挤甚至灭绝,这一过程被称为“竞争性排斥”。
例如,20世纪50年代,美国生态学家高斯做了著名的巴拉米西胶虫实验,证明当两种原生动物生活在同一瓶中、食物单一时,总有一种会被淘汰。这类似于我国淡水湖中大型外来鲢鱼入侵后,本土鲤鱼等滤食性鱼类被挤压生存空间。
生态位的动态变化
生态位并不是一成不变的,而是在环境、时间、甚至物种自身的生命周期阶段中不断调整。这种变化既可以发生在生态时间尺度(几年、几十年),也可以体现为个体发育过程中生态位的转换。
生态时间尺度的变化
在较短的时间内,环境变化(如气候、食物、竞争者的变化)会促使物种生态位发生动态调整。例如:
- 空间转移: 动物会因食物枯竭而迁移,如藏羚羊随季节更换栖息地。
- 食性改变: 干旱年份鸟类会增加杂食性植物的摄入比例来应对昆虫数量下降。
- 季节适应: 以秦岭金丝猴为例,夏季主要在树冠层觅食嫩叶和果实,冬季则转向树皮和芽苞,这种季节性生态位调整极大地提高了其生存弹性。
此外,有些植物可以通过改变根系深度,在不同季节利用不同层次的水资源,从而表现出“动态生态位宽度”。
生命周期中的生态位转换
许多动物在不同发育阶段拥有完全不同的生态位,这种现象被称为生态位转换。例如:
- 青蛙: 幼体蝌蚪期生活在水中,以浮游生物为主食;变为成体后,转为陆生、以昆虫为食。
- 鱼类: 许多鱼类幼体在河口或浅滩生活,成体则迁入深水区。
从图中可见,中华虎凤蝶卵、幼虫和蛹阶段活动范围极小,主要生活在地面或低矮植被上;但成虫阶段活动范围骤然扩大,在林间高空飞翔并寻找配偶与产卵地点。这种生态位转换有助于物种避开不同捕食者与竞争对手,从而最优化各阶段的生存策略。
植物也会发生生态位转换,例如一些杂草幼苗期需强光,成长期能耐阴;某些鱼类(如中华鲟)会在幼年时溯河进入淡水洄游,而成年后长时间生活于海洋。
生态位宽度

生态位宽度(Niche breadth)描述的是一个物种能够利用多少种类的资源及其环境适应范围的大小。一般来说,如果一个物种仅利用极少数类型的资源或很窄的环境,则其生态位宽度较窄,被称为专家型(特化物种)。反之,能在多种环境、利用多样资源的,则为通才型(泛化物种)。两者之间没有绝对优劣,各自拥有不同的生存策略与生态适应方式。
专家型物种的优势与劣势
专家型物种在特定环境下能展现极高的效率,通常在获取特定资源、对特定环境的适应和竞争中处于优势。例如:
中国的典型专家型物种:
- 大熊猫:99%的食物来源是竹子,消化系统高度特化,仅适应竹林生态系统,对竹子周期性枯死或减少极为敏感。
- 藏羚羊:专门适应青藏高原寒冷、缺氧的环境,生理结构和行为都适应高原,无法在低海拔地区存活。
- 白鱀豚:只生活在长江及其下属水体中的淡水环境,对环境变迁及污染极其敏感(目前已灭绝)。
通才型物种的适应策略
通才型物种相较于专家型,更能适应多变和复杂的环境。例如:
中国的典型通才型物种:
- 猪獾:杂食性很强,食物包括植物根茎、水果、昆虫、小型脊椎动物等,可以在田野、林地、村庄附近生存。
- 乌鸦:不仅适应城市、农村与郊野,还能利用各种食物,如垃圾、作物、动物尸体等,被称为“生态清道夫”。
- 麻雀:适应力极强,能够栖息在乡村、城市、农田和林缘等多种环境,筑巢和觅食策略多样。
通才型物种的优点:
- 当外部环境突变或资源类型发生更替时,依靠对多种资源的利用,存活概率更高。
- 可以快速扩散到新环境,对人类活动或自然干扰有较强恢复力。
潜在劣势:
- 在单一资源极丰富或某种环境高度稳定时,不如专家型“专一、高效”。
下面展示了泛化物种与特化物种在不同资源上的利用效率对比:
可以看出,大熊猫只在少数几类资源(主要是竹子的不同部分)上有较高利用效率,其它类型资源基本不利用(“-”),而猪獾则能对多数资源都进行中等程度利用。
生态位宽度的动态性与进化意义
生态位宽度不仅受遗传与进化的影响,还可能随着环境、种群压力甚至物种自身历史阶段而调整。有些物种在资源丰富时趋向于专一化,而在资源匮乏或竞争压力大时,会扩展自身生态位(即“生态位扩张”)。反之,长时间的环境稳定也可能催生新一轮的专一化进化。
环境决定最优策略
在资源长期稳定、竞争压力明确且主要资源充裕的情况下,专家型(特化)策略有优势,物种可以深化对特定资源的开发而减少竞争。但在环境波动剧烈、资源种类多变,尤其是人为干扰、气候异常或生态系统易于扰动的情况下,通才型(泛化)策略更容易保证物种的生存和扩散,因此这样的种群在新环境和生态位中更具备“先行者”优势。
生态位宽度也常用于预测外来物种入侵(通常泛化物种入侵风险更高)、保护濒危物种(专家型物种受威胁更大),以及应对气候变化过程中的生态稳定性分析,是生态学研究的经典主题之一。
最优觅食理论与斑块环境利用
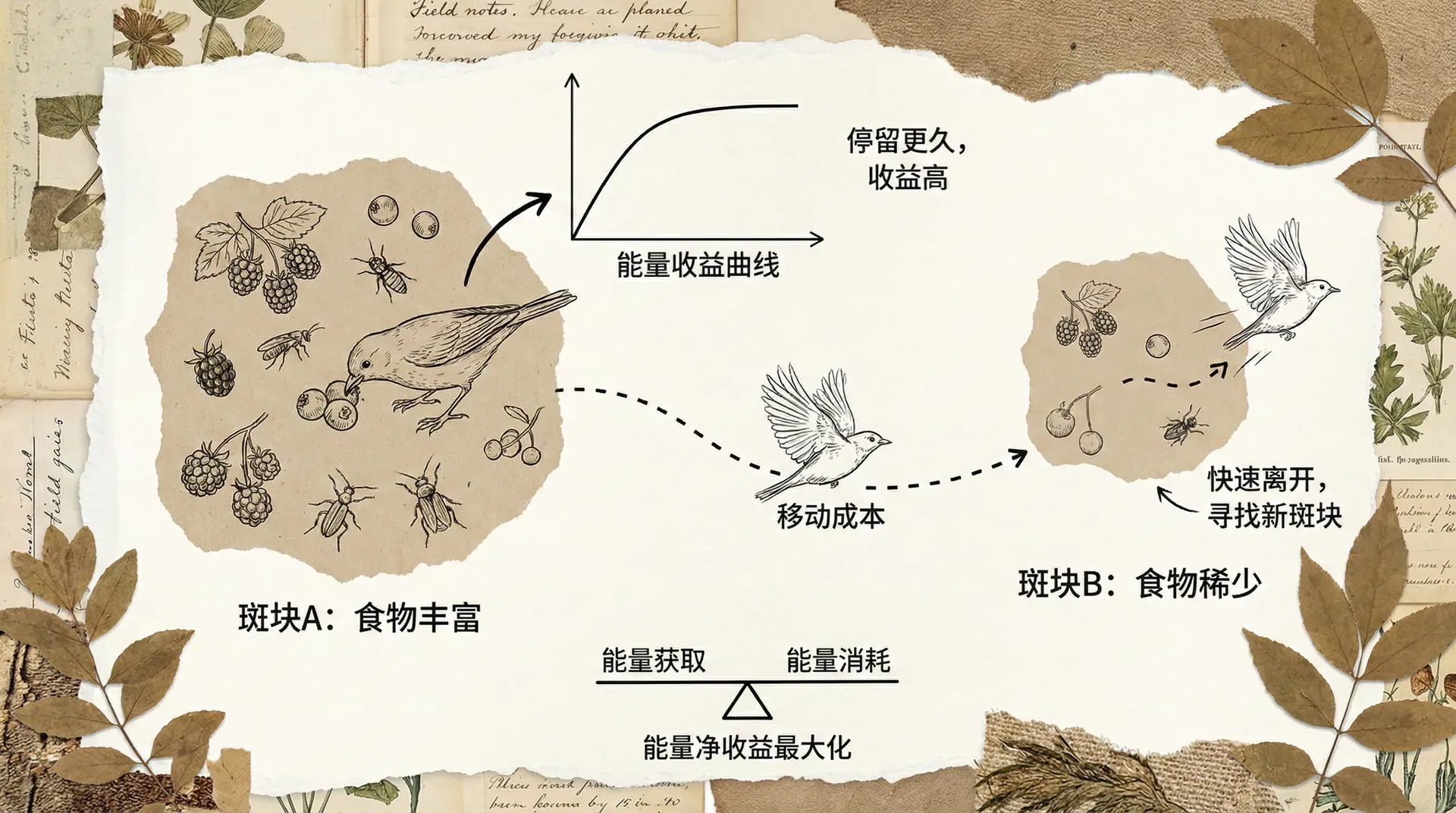
在自然界中,食物和其他资源通常呈现出斑块状分布,这意味着某一区域食物较为丰富,而另一区域则较为贫乏。动物在这样的环境中觅食,并非简单地“碰到什么吃什么”,而是要在不同斑块之间做出权衡决策。这一过程核心是“最优觅食理论”(Optimal Foraging Theory, OFT),它旨在解释动物如何以最小的成本获得最大的能量收益。
动物必须权衡“搜索时间”(找到各种食物的时间)与“处理时间”(摄取和消化食物所需的时间),并根据环境变化动态调整觅食策略。最优觅食模型假定动物会在资源丰富但开采速度变慢(搜寻和处理成本变高)时切换斑块,避免在某一地点耗费过多时间。
以梅花鹿在森林-草地镶嵌景观中觅食为例,它面对的决策包括:
- 当前斑块的食物质量 —— 即剩下的食物数量和易获得性;
- 移动到下一个斑块的时间与能量成本 —— 包含途中能量消耗、遭遇风险的概率;
- 新斑块的预期能量回报 —— 是否值得迁移以获取更多或更优质的食物。
这种决策模式其实是生态学广泛关注的“边际价值定理”(Marginal Value Theorem)的应用,强调动物会在当前斑块的收获边际收益降到等同或低于其他斑块的平均收益时,才会决定离开并转移。
从上图可以看出,梅花鹿如果只专注于极少种类的食物,虽然处理时间较低,但寻找这些目标的时间非常长,导致总成本很高。反之,如果什么都吃,虽然搜索容易(哪都有可食用对象),但各种食物的消化和处理效率不同,处理成本大幅提高。经过综合权衡,选择4种食物类型时,总成本最低,这就是所谓“最优觅食宽度”。这一点不仅对梅花鹿成立,对许多杂食动物和捕食动物均适用,是动物行为生态学中的经典理论。
实际上,最优觅食理论还可以解释捕食者选择猎物、鸟类挑选种子以及蜜蜂采花等现象,帮助我们理解生态系统中能量流动的微观机制。
竞争压缩假说
当生态系统中新竞争者加入时,资源压力陡然上升,原有物种不得不重新分配其生态位。生态学家提出的“竞争压缩假说”正是描述这种动态变化的理论。根据该假说:
当竞争压力加大时,物种会缩小其栖息地或活动空间范围(空间生态位收缩),但可能会扩大对食物种类的利用范围(营养生态位扩展)以维持生存。
例如,一条街只有一家餐厅时,可以做生意到很远的区域、只专注几道菜。若同街餐厅陡增,顾客被分流,这家餐厅会收缩外送距离,同时不断开发新菜式以吸引不同口味的顾客。
在鸟类群落研究中,科学家发现:某些鸟类面对新入侵者时,其原来广泛分布于多种林型与草地,但竞争加剧后活动范围减小,取而代之的是食性变杂、开始尝试原先不太选择的果实或昆虫。
从图中可以直观地看到,栖息地类型利用范围无论是广度还是强度都随着竞争加剧而缩小。这种竞争压缩现象在鱼类、昆虫乃至植物中都有类似案例,表现为生态位各维度的动态调整,是群落结构变迁的重要机制之一。理解这一理论有助于保护生物学、外来物种管理和恢复生态学等领域的实践工作。
总结
生态位理论揭示了生物如何通过复杂而微妙的分工和调整,确保每个物种在自然界中的一席之地。无论是在苛刻的荒野、激烈竞争的群落,还是高度人类化的环境中,生物都在不断寻求最优生存路径。这不仅关乎每个物种的“独特职位”,也影响整个生态系统的稳定和动态平衡。
掌握生态位理论,不仅有助于我们保护珍稀物种,预防外来入侵,还能指导我们优化农业和城市生态系统,实现人与自然的和谐共处。面对全球变暖、栖息地破碎化等重大生态挑战,生态位理论为生态学家、保护工作者和决策者提供了科学的分析框架:我们可以据此预测物种命运,评估生态风险,制定多层次的保护和管理举措。
正如生态学家奥德姆所说,认识了每个物种的“职业”,我们就有能力进行整个生态系统的统筹规划,实现人与自然社会真正的可持续发展。