系统发育学
如果我们要研究你的家族历史,我们会查看家谱,梳理祖父母、曾祖父母的身份,以及你和堂兄弟姐妹的亲缘关系。在生物学中,科学家们也在进行着类似的工作,不过我们研究的不仅仅是人类的家族,而是自然界中各种不同生物之间的“亲戚关系”。这个庞大的研究领域,就是“系统发育学”。它的核心目标,就是揭示物种与物种之间的演化联系和进化顺序,帮助我们搞清楚今天生物多样性的由来。
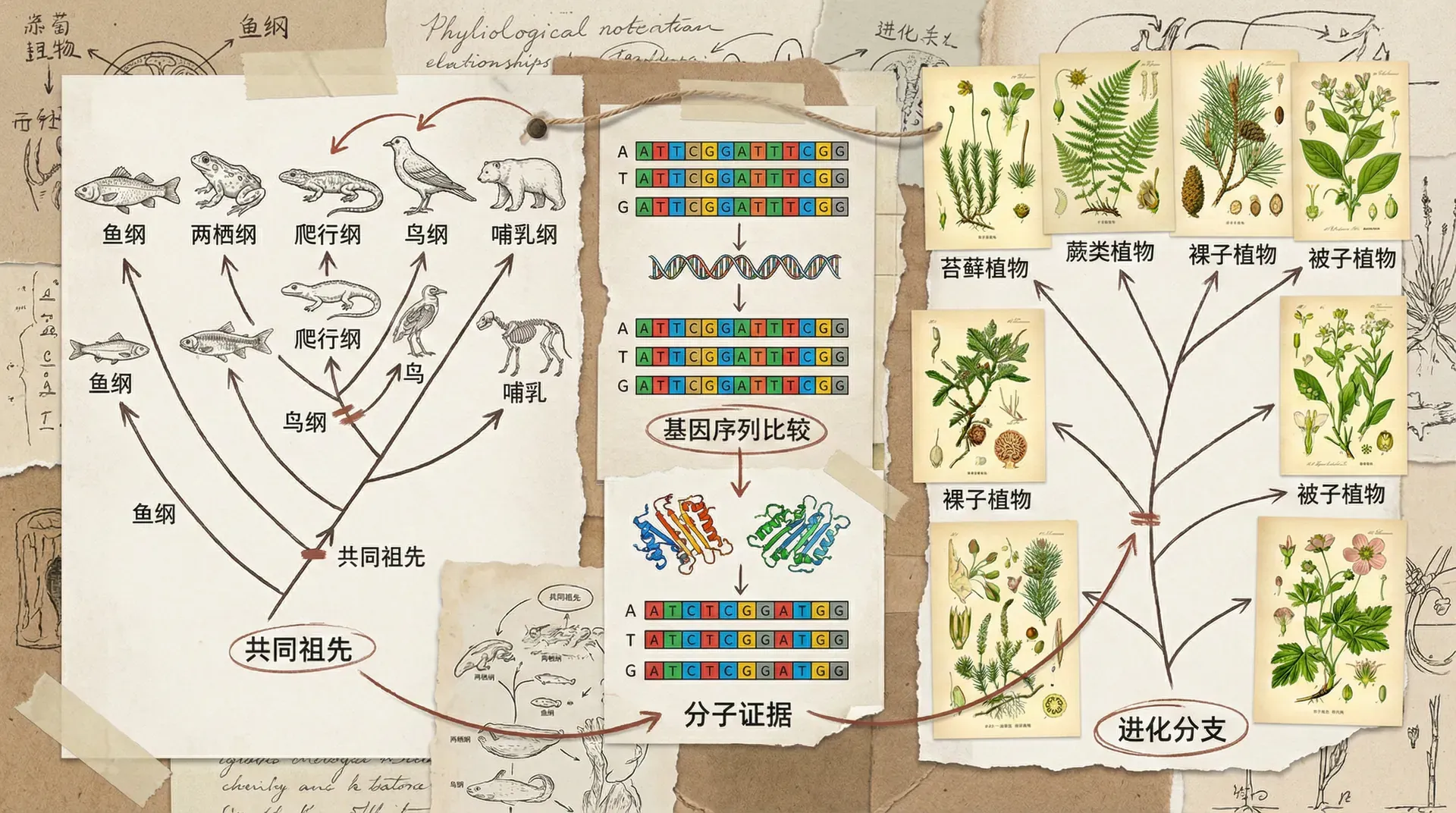
什么是系统发育树
系统发育树(Phylogenetic Tree)可以看作是自然界的“家谱树”或“族谱”。它用树状结构展示不同生物是如何从一个共同祖先逐步分化出来的。树干代表着那些久远年代的祖先生物,每一个分枝对应着进化过程中一次又一次的“分家”,直到树枝的末端,出现了现存的多样物种。通过解读系统发育树,我们能够了解物种的亲缘远近,推断它们共同祖先的面貌,还可以追溯它们各自演化出独特特征的过程。
系统发育学不仅仅是翻译或整理已有的国外文献,更是一项需要深入考察和推理、融汇多学科证据的科学探索。生物学家就像侦探一样,通过观察现存物种的形态结构、分子数据,甚至生活习性,综合分析,逐步拼凑出“演化历史的拼图”。
科学家在构建系统发育树时,十分重视“共有衍生特征”(synapomorphy)。这些特征是某一演化分支上的生物共同拥有,但其近亲或祖先没有的新特征。比如,大熊猫拥有著名的“伪拇指”——其实就是腕骨特化变形而成,帮助它抓握竹子,这在其他熊科成员中并不存在。这样一项独特性状就成了辨别大熊猫分支、理解其演化历程的“证据链”之一。此外,随着分子生物学的发展,科学家可以通过分析不同生物的DNA和蛋白质序列,进一步精确地绘制和校准系统发育树,使判断更为科学和权威。
地理如何塑造生物演化
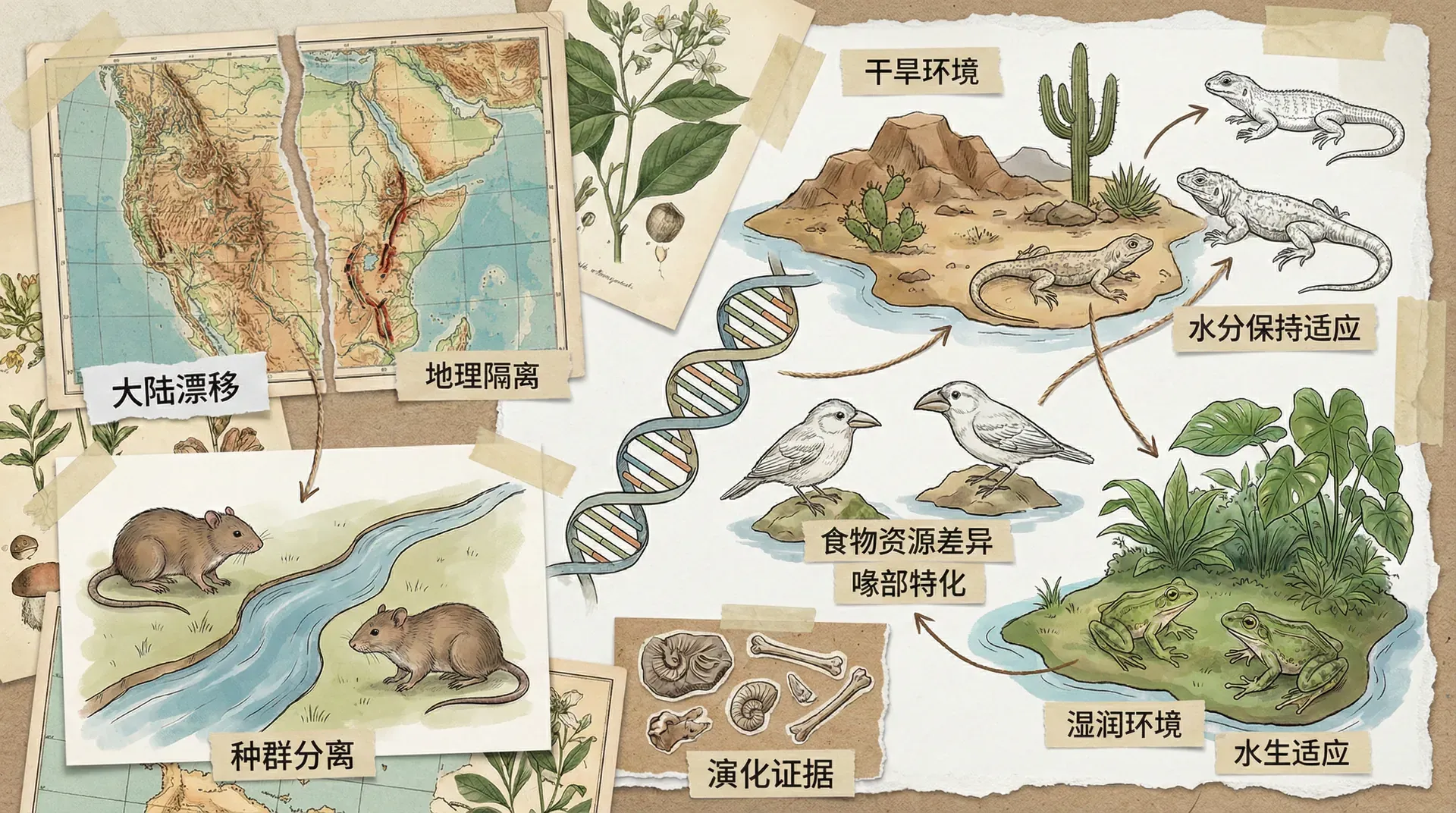
在研究生物演化时,我们不得不提到地球环境的巨大影响。地球并不是一成不变的:大陆漂移、火山喷发、冰川消融,乃至河流改道、山脉隆起……所有这些地质变化,都可以改变生物的生存环境,把原本生活在一起的物种种群“物理隔离”开来。就好像一面新的围墙,把原本亲密的邻居强行分开,从此各自面对不同的生活环境。这些孤立种群在不同的环境里适应进化,久而久之就可能分化为新物种,推动了生物多样性的涌现。
约在5000万年前,印度板块与亚洲板块碰撞,造就了世界屋脊——青藏高原。这个过程中,原本分布广泛、连为一体的动植物种群被新形成的高原和山脉隔断,分别在高原和平原两个生态系统中生存。高原上的空气变得稀薄、气温低、紫外线增强、氧含量减少,这对动植物来说,是非常严酷的挑战。只有能够适应这些极端环境的生物,才能留下后代,最终演化出许多高原特有的“新物种”。中国西部特有的旱獭、高原鼠兔、雪莲等,都是青藏高原隆起下的“演化明星”。
地理隔离仿佛为生物演化按下了“分支”按钮——原本同宗同源的种群,在天堑的两端开启了各自迥异的进化历程。正是这种分分合合、不断分化与适应的演化机制,让生物界变得五彩斑斓。
我们不妨来看一个数据化的例证:青藏高原地区植物多样性随海拔变化的规律
从上图可以看出,青藏高原的植物物种数量在海拔3000米左右达到峰值。这一带气候温和,水源充足,适合各类植物生长。随着海拔进一步升高,气候变冷、空气中氧气含量显著下降,紫外线更为强烈,这些高原环境因素使得能够生存的物种大大减少。由此形成的梯度分布,正是地理和环境因素对生物多样性“筛选”作用的生动写照。此外,这种山地生物多样性的“中高峰、两头低”的趋势,在许多大陆高原和山系中都有广泛的体现,展现了自然界在分化、隔离和适应中的复杂性与多样性。
从现在推测过去
我们无法回到几百万年前,亲眼见证生物祖先的模样与演化过程,但这并不妨碍我们通过研究现存物种,推断它们祖先可能拥有的特征。这就像通过对多个表兄弟姐妹的外貌比对,可以反推出他们共同祖父母的形象轮廓一样。科学家们把这种方法称为“比较法”或“系统发育推断”,是演化生物学中非常核心的研究思路。
在中国分布的金丝猴主要有川金丝猴、滇金丝猴、黔金丝猴等不同亚种,它们各自生活在西南地区不同纬度和海拔的山地森林中。不同生态环境塑造了它们显著的形态和生态差异——有的体型庞大,有的毛发浓密且色彩各异,有的则善于攀爬陡峭山坡,有的适应于寒冷高原甚至在雪地中活动。
这些差异绝非偶然。科学家们系统调查和比对后发现,生活在纬度和海拔更高、环境更寒冷的金丝猴,往往体型更大、毛发更长更密。这是经典“贝格曼定律”和“艾伦定律”的生动体现:体型越大、四肢越短,有利于减少热量的散失,更加适应寒冷气候。而相对温暖潮湿、海拔较低地区的金丝猴则普遍体型较小,毛发颜色和密度也发生相应变化。
此外,科学家还会结合遗传信息(如DNA序列),以及化石证据,进一步追溯金丝猴家族的演化分支和年代。通过比较现存亚种的形态、分布和遗传距离,可以推测它们最近的共同祖先生活在什么区域、具体有什么样的生态适应。比如,有研究推测,多种金丝猴亚种大约在百万年前因地理隔离和气候变化而分化开来,祖先可能体型居中,毛色偏深,分布在横断山区一带,随后随着青藏高原的隆升与气候波动,逐渐扩散到不同的山地生态带。
下方观察金丝猴体型与纬度之间的数据关系:
从图中可以清晰地看到,随着纬度升高(气温降低),金丝猴的平均体重也随之增加。这条“北大南小”的趋势不仅揭示了环境条件对金丝猴形态的深刻影响,也为研究者提供了逆向推测祖先特征的可靠线索。这背后反映出生物体如何在不同地理与气候环境下,通过自然选择,不断调整自身特征以适应新的生态挑战。而“现存物种的比较”,正是我们一步步揭开历史演化面纱的“时光望远镜”。
形态背后的生态故事
生物的身体结构绝非偶然产物,而是漫长演化与自然选择下的成果,深刻反映出它们在生态环境和生活方式中的不断适应。不同生物的外形,就如同不同运动员根据项目锻炼出最适合自身需求的体型,是其“生态职位”(生态位)的直观映射。一个物种之所以呈现现在的样貌,背后离不开在求生、繁殖、觅食等多方面的“设计优化”。
雉类鸟的腿长和生态适应
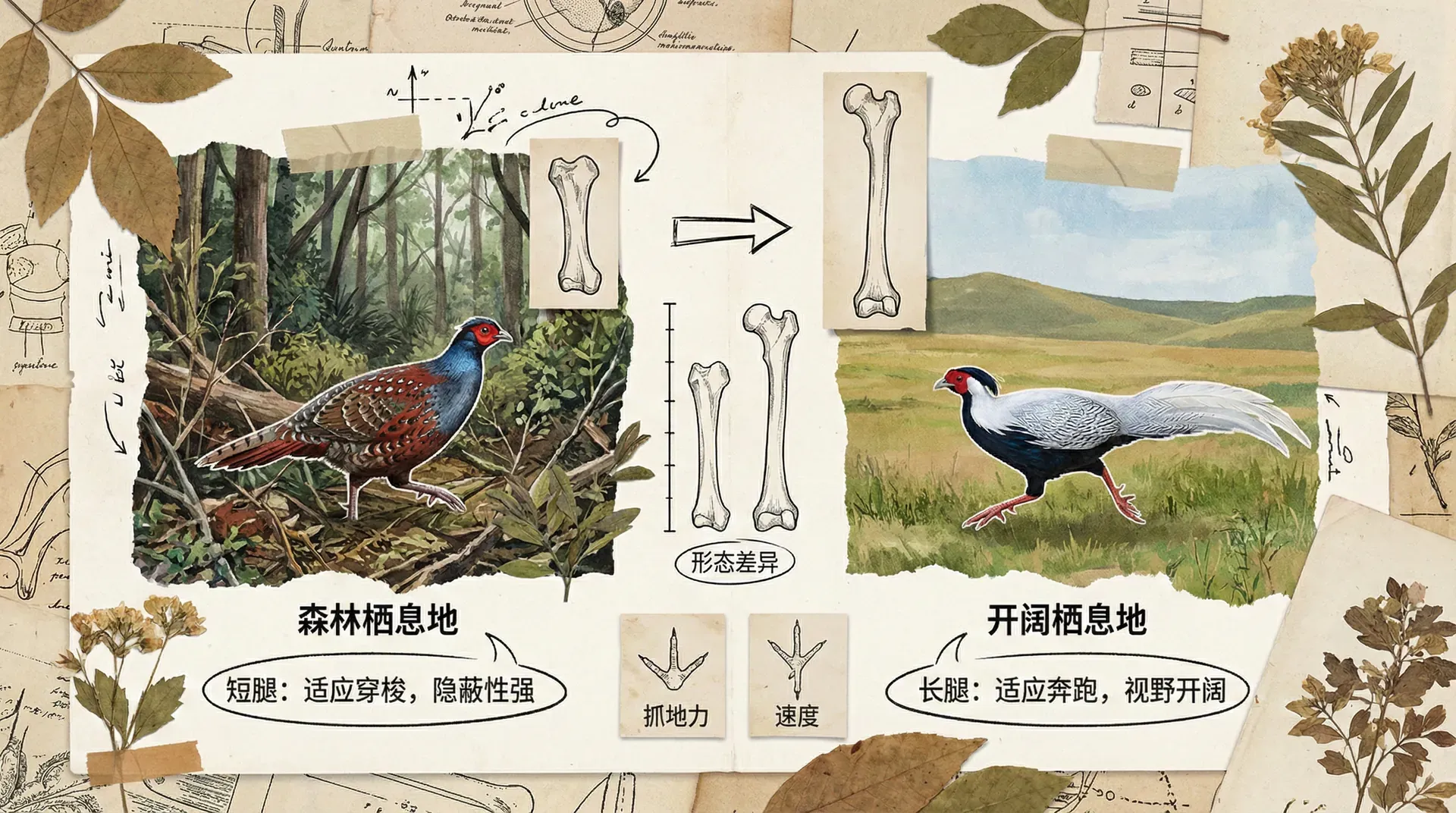
以中国山地森林中的雉类鸟为例,它们形态各异、生态分化明显:部分,如红腹锦鸡(又叫金鸡),偏好阳光充足、开阔的草地;而血雉、绿尾虹雉等则常活跃于幽暗密林。不同的栖息环境,造就了两类雉鸟在腿长上的显著差别。草地雉类由于需要疾跑、跳跃逃避捕食者,演化出较长且强健的双腿,地面疾走优于飞翔。而密林雉类则腿短结实,适应穿行于障碍丛生的林地,更善于隐蔽和穿插。
下方表格汇总雉类鸟栖息地与腿型差异:
形态特征如同生物的“身份证”,帮助我们识别物种,还能洞察其生态习性与生活环境的秘密。
为了更直观地了解环境与形态的关系,我们来看一组数据:
可以看到,森林、草原和林缘雉类在腿长与环境开阔度上的数据分布各有侧重。密林中的血雉腿短适合隐蔽,草原上的红腹锦鸡腿最长适合奔跑,林缘的白鹇则介于二者之间。这一系列演化适应,使不同物种精准占据各自的生态空间。
更多“外形—生态”案例
实际上,“腿长—生活环境”只是众多形态-功能关系的一例。我们还可以举出如下有趣的实例:
此外,鸟类的翅型和飞羽结构同样与飞行方式紧密相关。例如:栖息林间的松鸦、画眉,拥有较圆短的翅膀,便于在林间穿梭急转;而如金雕、鹗则拥有长而狭的翅膀,适合滑翔、长距离飞行。
这种“形态-生态”一一对应关系不仅帮助我们理解现生物种多样化的奥秘,还极具科学意义——古生物学家在发掘化石时,常通过骨骼比例等形态特征,推断古动物的生态环境、活动方式,甚至可能的取食行为。例如,恐龙的股骨与胫骨比例可判断其奔跑能力,牙齿结构则透露食性。
随着科技进步,现代研究还会将生态学、气候学和分子生物学等多领域数据整合在一起,比如收集几十种鸟类的腿长、翅膀类型、纬度分布、食物来源等,利用统计学和系统发育树等工具,揭示隐藏于表型背后的演化规律。这些例子让我们看到,动物外形背后不仅有生活环境的烙印,更蕴藏着物种演化的壮美故事。
总结
系统发育学为我们提供了一种全新的视角来理解生命世界。通过构建系统发育树,我们能够追溯物种的演化历程;通过研究地理隔离事件,我们能够理解生物多样性的形成机制;通过比较分析,我们能够从现存物种推测祖先的特征;通过形态生态学,我们能够解读生物形态背后的生态意义。
这些方法相互结合,让我们能够像阅读一本书一样,逐渐读懂地球生命演化的历史。虽然很多物种已经灭绝,但它们留下的痕迹——无论是化石、DNA序列,还是现存近缘种的特征——都在向我们诉说着那些逝去的故事。