走进生物心理学
人为什么会恐惧?为什么有些人天生乐观,有些人却容易陷入低落?为什么双胞胎在相似的成长环境下,性格却可能截然不同?这些问题表面上是心理学的范畴,但真正的答案却深藏在大脑的神经回路、基因的碱基序列以及数百万年的进化历程之中。
生物心理学正是那座连接生物与心理的桥梁。它研究的不是抽象的“灵魂”或“意志”,而是神经元的放电模式、激素的浓度变化、基因在不同环境下的表达方式。它的核心假设是:我们所有的思想、情感和行为,都对应着大脑中某种可被测量的生理状态。
这个学科让我们直面三个根本性的问题。心理与大脑之间究竟存在怎样的关系?我们的行为究竟有多少来自遗传,又有多少由经历塑造?在探索这些问题的过程中,科学的边界在哪里,伦理的底线又在哪里?
生物心理学并不是要把人简化为一堆化学反应,而是从生理机制、个体发育、进化历史和适应功能这四个维度,去理解行为为何如此、从何而来。
两个永恒的哲学问题
在人类所有的追问之中,有两个问题尤为根本,也尤为困难。它们一个指向宇宙本身的存在,另一个指向意识与物质之间的关系。理解这两个问题,是走进生物心理学之前不可绕过的思想准备。
宇宙为何存在
德国哲学家莱布尼茨在1714年提出过一个令人头晕的问题:“为什么存在某些东西,而不是什么都没有?”从直觉上来说,什么都不存在似乎才是更“自然”的状态。然而宇宙就在这里,星系、行星、生命,一切都真实地存在着。这是怎么发生的?
暂且搁置这个最终极的问题,还有一个稍微容易探讨的问题:既然宇宙存在,为什么是现在这个样子?我们的宇宙包含质子、中子、电子,它们各自具有精确的质量与电荷,四种基本力——引力、电磁力、强核力、弱核力——以特定的强度比例共存。这些数值哪怕有一点点偏差,宇宙的面貌就会完全不同。
从1980年代起,弦理论家们试图用数学证明这是宇宙唯一可能的形态。但结果令人意外:方程给出的可能宇宙数量,是1后面跟着500个零那么多。在所有这些可能的宇宙中,能够支持生命存在的极为罕见。
引力若稍弱,物质无法凝聚成恒星和行星;引力若稍强,恒星燃烧殆尽的速度快过生命诞生所需的时间。两种力的精确比例,使氦与铍在恒星内部能共振融合成碳——而碳是一切已知生命形式的基础元素。任何一种力改变不到1%,宇宙中几乎就不会有碳。
水的存在同样令人称奇。大多数轻分子在常温下是气体,而水分子中两个氢离子以104.5度角结合,造成分子两端微弱的正负电荷差,足以让水分子彼此吸引,在常温下保持液态。吸引力若稍弱,水就全是蒸汽;若稍强,则永远是冰。正是这种微妙的平衡,使地球上的液态水得以存在,也使生命得以延续。
简而言之,宇宙本可以以无数种形态存在,而在几乎所有的那些形态中,生命都不会出现。
意识的困难问题
第二个深刻的问题,哲学家戴维·查默斯将其称为“困难问题”:在这个由物质与能量构成的宇宙里,为什么会有意识这种东西?
我们可以描述神经元如何放电、突触如何传递化学信号、大脑的哪个区域在视觉刺激下活跃。但这些描述都停留在物理层面。从“大脑的某个部位产生了电活动”到“我看见了红色”,中间隔着一道至今无人能跨越的鸿沟。为什么物质的运动会产生主观体验?这就是心理与大脑的关系问题,也是整个生物心理学试图靠近的核心谜题。
目前还没有人能给出令人信服的答案。有人主张彻底放弃“意识”这个概念,但这更像是回避问题。查默斯等人提出将意识视为物质的基本属性,就像质量和电荷一样无需还原解释。然而这个答案并不令人满意——质量和电荷在宇宙中无处不在,而意识只出现在某些神经系统的特定状态下,昏迷时没有,深度无梦睡眠时也没有。
把某样东西称为“基本属性”,并不意味着它真的没有原因,而是意味着我们目前还不知道原因,或者暂时放弃了寻找原因。意识问题是否能被科学最终解答,至今仍是未知数。
这两个问题——宇宙为何存在、意识为何出现——共同构成了生物心理学研究的哲学背景。前者提醒我们,生命本身就是一个极不寻常的存在;后者则提醒我们,即便大脑的生理机制被完整描绘出来,关于“体验”的问题或许依然悬而未决。
生物心理学的本质
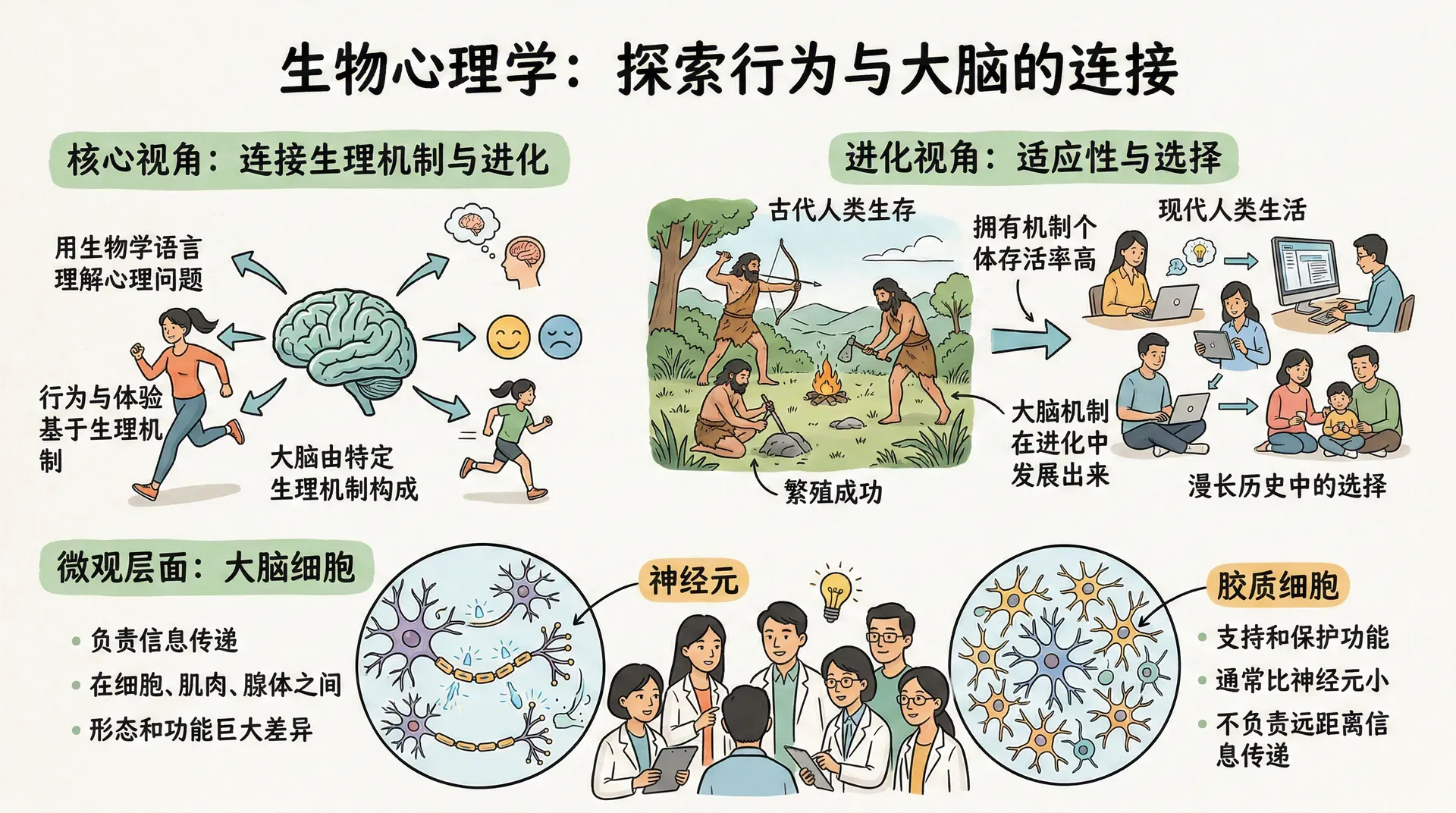
生物心理学是研究行为与体验的生理机制、进化历史及发展过程的科学,与心理生物学、生理心理学、行为神经科学等学科高度重叠。这些名称侧重点略有不同,但共同指向一个核心目标:用生物学的语言来理解心理学的问题。
这门学科的基本立场是,人类之所以会以某种方式思考和行动,是因为大脑中存在相应的生理机制,而这些机制之所以进化出来,是因为在漫长的历史中,拥有这些机制的个体比其他个体存活率更高、繁殖更成功。在微观层面,大脑由两类细胞构成:神经元负责在细胞之间以及向肌肉和腺体传递信息,它们在形态和功能上差异巨大;胶质细胞通常比神经元小,承担支持和保护功能,但不负责远距离的信息传递。
生物心理学不是把人简化为机器,而是在承认心理现象真实存在的前提下,去追问这些现象的生物学基础是什么,以及它们是如何在进化过程中形成的。
行为的四种生物学解释
日常生活中,我们解释行为的方式通常是围绕意图展开的——“他这样做是因为他想要……”“她那样做是因为她决定了……”。但这类解释有一个隐含的前提:行为者是有意识地做出选择的。现实情况并不总是如此。一只刚出生几个月的候鸟第一次向南飞,它并不知道自己为什么要这样做;受惊时起鸡皮疙瘩的人,也不是刻意控制毛囊肌肉的。
生物学对行为的解释分为四个层次,每个层次回答的是不同类型的“为什么”。
生理解释
生理解释关注的是此时此刻大脑和身体的运作方式,即行为在神经和生化层面的即时机制。它追问的是:当某个行为发生时,身体里发生了什么?睾酮浓度升高时,动物变得更具攻击性,原因在于睾酮作用于大脑特定区域的受体,改变了神经元的活动模式。这就是一个生理解释。生理解释不涉及行为的起源或功能,只描述机制本身。
个体发育解释
个体发育解释追问的是行为或能力是如何随个体成长而形成的。“个体发育”来自希腊语,意指“存在的起源”,在这里指的是一个生命体从受精卵到成年的发展过程。这类解释关注基因、营养、早期经验及其相互作用如何共同塑造了当下的行为能力。
抑制冲动的能力就是一个典型。这种能力在幼儿期几乎不存在,到青少年期才逐渐发展起来,原因在于大脑前额叶皮层的成熟过程要一直延续到二十多岁。这不仅仅是教育的结果,更是大脑结构发育的时间表所决定的。
进化解释
进化解释重构了一种行为或结构的演化历史,追问的是它从哪里来。动物的大多数特征都是祖先物种特征的修改版本。黑猩猩偶尔会用树枝取食白蚁,这一行为在灵长类中有着古老的根源;人类对工具的精细操控,则是在这个基础上经过数百万年的选择压力逐步演化而来的。
进化解释也能解释那些看起来已经“没用”的特征。人在受到惊吓时会起鸡皮疙瘩,这在人类身上几乎毫无意义——我们的体毛太短,竖起来既不能保暖,也不能让自己看起来更大。但在大多数哺乳动物中,全身毛发竖起可以让动物显得更大、更具威胁性,是一种有效的防御信号。人类保留了这个反射,只是它已经失去了原本的功能。
功能解释
功能解释追问的是某个特征或行为为什么会被自然选择保留下来,它带来了什么适应性优势。许多动物的体色与栖息环境高度吻合,这一现象的功能解释是:体色越接近背景,被天敌发现的概率就越低,生存率也就越高。
需要注意的是,功能解释描述的是历史上的优势,而不一定是今天的优势。某个基因在过去的环境中有利于繁殖,并不意味着它在当下的环境中依然如此。
这四种解释方式可以用鸟类鸣唱这一行为来加以整合:
这四种解释层次并不互相矛盾,而是从不同角度共同描述同一个行为。真正理解一种行为,需要同时考虑它的即时机制、发展过程、进化历史和适应功能。
生物心理学的研究方向
生物心理学并非一个封闭的实验室学科,它的研究成果和应用遍及医学、教育、临床治疗等多个领域。了解这个学科的研究方向,有助于理解它为什么值得深入学习。
研究职位通常要求心理学、生物学或神经科学方向的博士学位,从事的工作包括设计实验、分析脑影像数据、研究特定神经系统疾病的机制等。许多研究者在高校兼顾教学与科研,也有人在医学研究机构或制药企业的实验室专注于应用研究。中国近年来持续加大对脑科学的投入,“中国脑计划”的推进带动了相关领域人才需求的增长,北京、上海等地的神经科学研究中心对具备扎实生物心理学背景的研究人员有持续的需求。
实践应用方向则覆盖面更广。神经科医生诊断和治疗脑部疾病,精神科医生通过药物或心理手段干预情绪和行为障碍,临床心理学家在评估和治疗心理问题时需要具备识别神经系统异常的能力。学校心理学家在判断学生是否存在注意缺陷、学习障碍等问题时,同样需要理解行为背后的神经发育机制。
无论从事哪个方向,生物心理学的学习都要求具备跨学科的思维方式——既能理解神经元层面的微观机制,又能从行为和体验的角度来解读这些机制的意义。
遗传学与行为
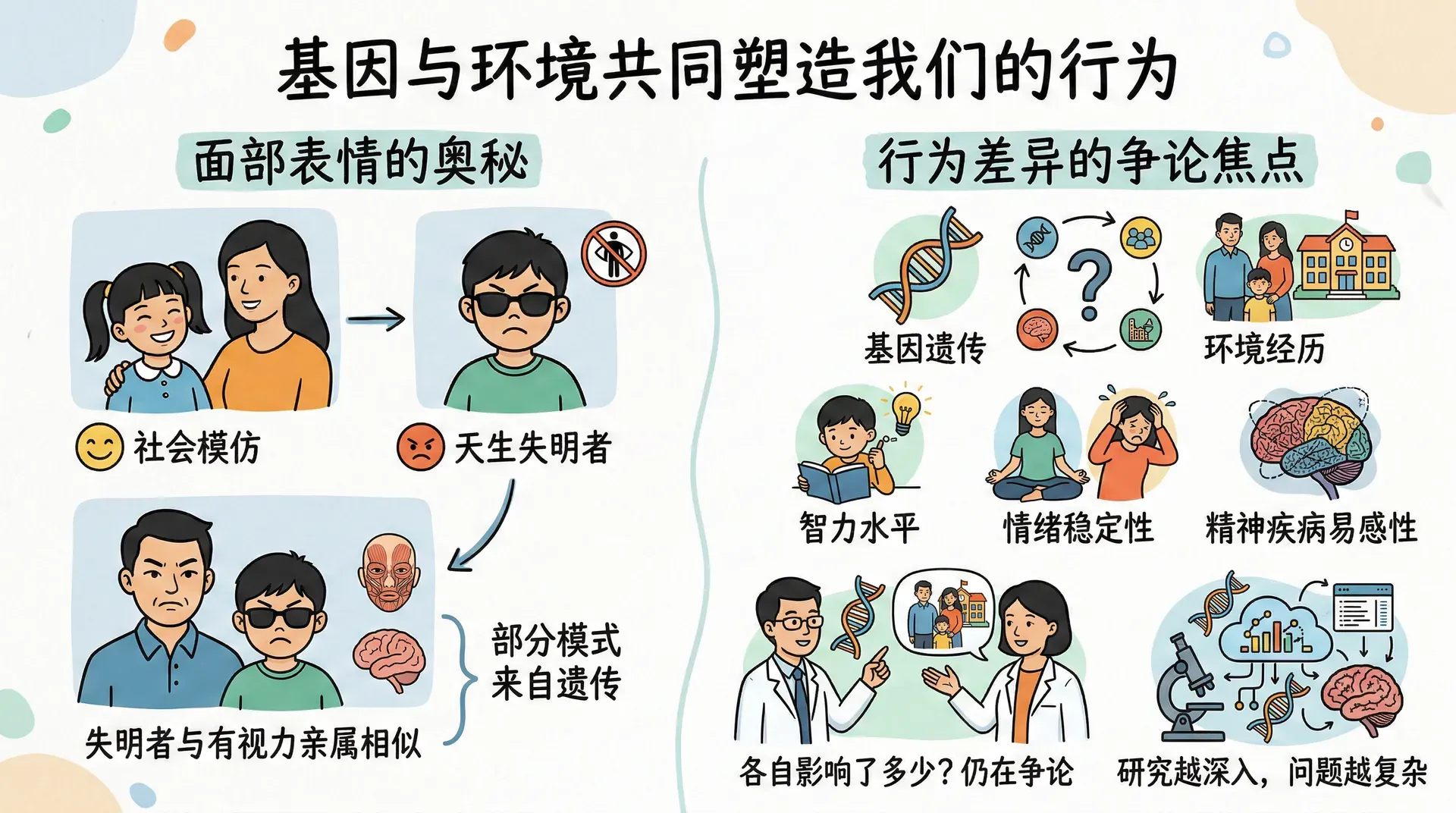
每个人的行为都同时受到基因和环境的影响。这个说法听起来不像是什么新鲜观点,但当我们试图弄清楚两者各自贡献了多少时,问题就变得复杂起来。
面部表情提供了一个直观的切入点。我们的表情当然受到社会环境的影响——开心时笑、难受时皱眉,这些与成长经历密切相关。但遗传的作用是否也在其中?研究者对天生失明的人进行了观察,这些人从未见过别人的表情,没有通过模仿学习的机会。结果发现,他们在高兴、悲伤、愤怒等情绪下的面部肌肉运动,与有视力的亲属高度相似。这说明某些面部表情的模式是写在基因里的,而不完全依赖于后天的观察和学习。
然而,一旦我们超越“遗传和环境都有影响”这个宽泛的结论,去追问“各自影响了多少”,分歧就会出现。人与人之间在智力水平、情绪稳定性、某些精神疾病的易感性方面存在明显差异,这些差异究竟主要来自基因还是成长环境,至今仍是争论的焦点。遗传学本身也在不断发展,研究越深入,问题往往越复杂。
孟德尔与遗传的基本规律
19世纪之前,科学界普遍认为遗传是一种混合过程,后代的性状是父母双方性状的中间结果,就像将两种颜色的墨水混合会得到第三种颜色。奥地利修道士格雷戈尔·孟德尔通过长达数年的豌豆杂交实验,打破了这个直觉上合理但实际错误的假设。
孟德尔发现,遗传是通过离散的单位进行的,这些单位在世代传递中保持结构完整——后来被称为基因。基因通常成对出现,因为它们沿着成对的染色体排列。唯一的例外是雄性哺乳动物的性染色体:X染色体和Y染色体携带的基因不同,它们不是真正意义上的“一对”。
从分子层面来看,基因是DNA分子上的一段序列。DNA由四种碱基——腺嘌呤、鸟嘌呤、胞嘧啶、胸腺嘧啶——按照特定顺序排列而成。这个顺序决定了信使RNA的碱基序列,进而决定了蛋白质的氨基酸排列。蛋白质是身体结构的组成成分,也是驱动各种生化反应的酶。整个过程就是“DNA→RNA→蛋白质”这条信息流。
仅用四种碱基的排列组合,就能编码出构成人体所需的数万种蛋白质,以及调控这些蛋白质合成的所有指令。遗传信息系统的精密程度,远超我们通常的想象。
显性、隐性与基因表达的复杂性
在两条同源染色体上,同一位置的基因可能完全相同,也可能存在差异。两条染色体携带相同基因版本的个体称为纯合子,携带不同版本的则称为杂合子。
基因的效应强度各不相同。显性基因无论在纯合还是杂合状态下都会表现出明显效果;隐性基因只有在两条染色体上都携带该版本时才会显现。棕色眼睛对蓝色眼睛是显性的,所以一个携带一个棕眼基因、一个蓝眼基因的人,眼睛会是棕色。
但这类简单例子很容易让人产生误解,以为“有某个基因就一定会表现出对应的特征”,或者“一个特征只由一个基因决定”。实际上,眼睛的颜色受至少十个基因的共同影响,身高则涉及超过180个相关基因。很多基因的表达还取决于细胞类型、发育阶段和外部环境。影响行为的基因尤其如此,它们的效应往往是多基因共同作用的结果,且高度依赖于具体的发育和生活情境。
“某个基因决定某种行为”的说法在大多数情况下都是过度简化的。基因的作用是概率性的、条件性的,而不是决定性的。
性染色体与性别相关基因
位于性染色体上的基因称为性联基因,其他染色体上的基因则称为常染色体基因。在人类中,女性携带两条X染色体,男性携带一条X和一条Y。Y染色体很小,只编码约27种蛋白质,而X染色体编码约1500种蛋白质。因此,生物学上所说的性联基因,通常指的是X染色体连锁基因。
红绿色盲是X连锁隐性基因的经典案例。携带这个基因的男性,因为没有第二条X染色体来“覆盖”这个隐性效应,必然表现出色盲。女性则需要两条X染色体上都携带这个隐性基因才会色盲,概率远低于男性。这也解释了为何色盲患者中男性比例远高于女性。
与性联基因有所不同,性限基因存在于两性中,但只在某一性别中活跃。控制男性面部胡须生长或女性乳腺发育的基因,两性都有,但性激素决定了它们在哪个性别中得以表达。
基因的变化与表观遗传调控
基因并非一成不变的固定程序,它以多种方式发生变化,也可以在不改变DNA序列的前提下改变表达方式。理解这两个层面,对于认识行为的生物学基础至关重要。
基因变化最直接的形式是突变,即DNA分子中碱基序列的可遗传改变。仅仅将一个碱基替换为另一种碱基,就可能导致蛋白质的氨基酸序列发生变化,进而影响蛋白质的结构和功能。进化在数十亿年间持续筛选,大多数随机突变都是有害或中性的,但偶尔出现的有利突变可以带来深远影响。人类与黑猩猩在一个与语言发展相关的基因上只差两个碱基,正是这两处差异,在一定程度上修改了人类的大脑结构和发声器官,为语言能力的演化奠定了生理基础。
另一类突变是基因组特定片段的复制或缺失。在细胞分裂过程中,染色体的某一段可能被复制两次,也可能完全丢失。当这种变化发生在很小的片段上时,称为微复制或微缺失。研究发现,大脑发育相关基因的微复制和微缺失与精神分裂症的发病风险存在关联,这提示基因组的微小结构变异可能对神经系统的正常发育产生显著影响。
经验改变基因的方式
表观遗传学研究的是在DNA序列本身不发生改变的情况下,基因表达如何受到调控和改变。这一领域揭示了“遗传”与“经验”之间的边界远比人们通常认为的模糊。
身体的每一个细胞都含有完整的基因组,但不同类型的细胞中活跃的基因并不相同。肝细胞和神经元有着相同的DNA,却执行着截然不同的功能,原因就在于基因表达的差异。同一个基因,在胚胎早期、青春期和老年期的活跃程度也可能各不相同。更关键的是,外部经验同样可以改变基因的开关状态。
其机制涉及一种叫做组蛋白的蛋白质。DNA在细胞核中并非以舒展状态存在,而是缠绕在组蛋白上,形成紧密压缩的结构。要激活某个基因,DNA必须从组蛋白上部分展开,转录机器才能接触到对应的序列。组蛋白分子上有可以被化学修饰的“尾巴”,当乙酰基附着在这些尾巴上时,组蛋白对DNA的抓握会松弛,基因随之被激活;乙酰基被移除,组蛋白重新收紧,基因就被关闭。
DNA本身也可以被甲基化修饰。在基因起始区域的启动子序列上添加甲基,通常会压制该基因的表达;移除甲基则可以重新开启基因。这两种机制——组蛋白修饰和DNA甲基化——共同构成了基因表达调控的主要表观遗传手段。
孕期营养不良的母鼠所生的后代,会改变某些与能量代谢相关基因的表达方式,使身体更倾向于储存脂肪。这种适应在食物匮乏的环境中具有生存优势,但若后代生活在食物充足的环境中,则会大幅提高肥胖和心脏病的风险。
早年缺乏母亲照料的幼鼠,海马区某些基因的甲基化水平会持续偏高,导致这些基因在成年后长期处于低表达状态,使个体对情绪压力的应对能力下降。学习和记忆的形成同样依赖基因表达的变化——每次经历新事物,大脑中的化学环境就会发生变化,影响哪些基因被激活、哪些基因被抑制,从而在神经回路层面留下痕迹。
表观遗传的核心启示在于:你现在的经历不只影响你现在的状态,还可能通过改变基因的表达方式,对大脑功能产生持续的影响。“先天”与“后天”之间并没有一道清晰的界限。
遗传与环境对行为差异的影响
“唱歌能力是天生的还是后天练出来的?”这个问题本身就存在一个逻辑缺陷:没有基因,你甚至无法存活;没有成长环境,基因也无从表达。更有意义的问法是:在现实中观察到的个体差异,有多大比例来自遗传差异,又有多大比例来自环境差异?
测量遗传力的方法
研究者通过两类设计来估算遗传的贡献程度。第一类是双生子研究。同卵双胞胎携带完全相同的遗传物质,异卵双胞胎则平均共享约一半的基因,与普通兄弟姐妹无异。如果某种特征在同卵双胞胎之间的相似程度显著高于异卵双胞胎,就说明遗传在这个特征上发挥了重要作用。
值得注意的是,“同卵”并不等于“完全相同”。表观遗传的差异会使同一对双胞胎在基因表达上逐渐产生分歧,尤其是在经历不同的生活环境之后。但即便如此,遗传相似性本身仍然提供了有价值的比较基准。
第二类是收养研究。如果被收养的儿童在某些特征上更接近素未谋面的生物父母,而非一起生活多年的养父母,这种现象就很难用环境来解释,遗传的作用便呼之欲出。研究者还会考察“虚拟双胞胎”——年龄相近、同时被收养到同一个家庭的无血缘儿童。他们从婴儿期开始共享相同的成长环境,却没有任何遗传关联。若他们在某些行为特征上仍存在明显差异,就提示了遗传的独立影响。
遗传力是衡量某一特征在特定群体中个体差异有多少来自遗传差异的指标。研究表明,几乎所有被系统测量过的行为特征都具有可观的遗传力,涵盖孤独感体验、情绪稳定性、学习第二语言的速度乃至日常看电视的习惯。几乎唯一被发现没有显著遗传力的是宗教归属——信奉佛教还是基督教,主要由文化环境而非基因决定。
需要特别说明的是,遗传力的估计值并非普遍适用的常数,它取决于特定群体中遗传变异与环境变异的相对比例。在一个家庭教育风格差异极大的群体中,环境变异对行为差异的贡献会被放大,遗传力的估计值相应降低;在一个成长环境高度一致的群体中,遗传差异的影响就会更加突出。
遗传并不等于不可改变
高遗传力不意味着某种特征无法被改变,苯丙酮尿症提供了一个清晰的反例。这是一种遗传性代谢疾病,患者体内缺乏分解苯丙氨酸所需的酶,导致该氨基酸在血液和脑组织中积累,损害大脑发育,引发智力发育迟缓和情绪问题。苯丙酮尿症的遗传基础非常明确,但它完全可以通过饮食干预来管理。
中国在内的许多国家已将苯丙酮尿症筛查纳入新生儿常规检测项目。一旦确诊,医生会建议家长为婴儿安排严格限制苯丙氨酸摄入的特殊饮食。坚持这一饮食方案的儿童,大脑发育基本不受影响,可以正常成长。
苯丙酮尿症的饮食管理极为严格,需要回避肉类、鸡蛋、乳制品及含有阿斯巴甜的食品,同时补充专门配制的氨基酸配方。这对家庭来说是长期的挑战,但它有力地证明了一件事:可遗传的,不一定是不可干预的。
行为的进化基础
进化是群体中基因频率随世代变化的过程。这个定义有一点值得特别注意:进化本身并不等同于“改进”,它只是描述基因组成的变化,无论这种变化对物种的长远发展是否有利。
研究进化时,我们通常面对两类不同的问题。
-
第一类是“哪些物种从哪些祖先演化而来”,这依赖于化石记录和现存物种的比较解剖。
-
第二类是“进化通过什么机制运作”,这是一个关于过程本身的问题,答案要简洁得多:进化是自然选择作用于可遗传变异的必然结果。
自然选择的逻辑
进化的发生并非偶然,它是以下三个条件同时成立时的必然产物:
后代在遗传上与亲代相似,“像产生像”的规律使得有利特征能够跨代延续。
突变、基因重组和染色体片段的复制或缺失,不断向种群引入新的可遗传变异,这些变异会影响个体的生存与繁殖机会。
不同个体在繁殖上的成功程度存在差异,与更高繁殖成功率相关的基因,会在后代中逐渐变得更为普遍。
中国古代的育种实践早已对这一原理有所运用。历代养蚕人通过挑选吐丝量多的蚕蛾留种,逐代积累有利特征,这与达尔文所描述的“人工选择”在逻辑上完全一致。达尔文的贡献在于指出自然界中也存在类似的选择压力——在寻找食物、躲避天敌、吸引配偶方面更具优势的个体,其基因会在后代中扩散。在足够长的时间内,这个过程足以解释生命世界的广泛多样性。
关于进化的几个常见误解
进化论在日常理解中存在一些顽固的误区,有必要逐一厘清。
第一个误区是:不使用某种结构,它就会在进化中逐渐消失。这种想法源自拉马克的“获得性遗传”理论——他认为个体在生前获得或丧失的特征,可以直接遗传给后代。按照这个逻辑,经常锻炼的父亲会生出肌肉更发达的孩子,从不使用小脚趾的人其后代小脚趾会更小。然而,遗传物质是DNA,不是肌肉状态或脚趾的使用频率。一个人的体验无法直接改写精子或卵子中的基因序列,生物学家至今也没有发现支持拉马克式遗传的可靠证据。
第二个误区是:现代医学让人人都能活下去,人类已经停止了进化。这个论证混淆了“生存”与“繁殖”。进化的驱动力是繁殖成功率的差异,而非存活率。只要不同个体在生育数量上存在差异,且这些差异与基因有关,进化就在持续发生。
第三个误区是:进化等于“向更好的方向改变”。进化的衡量标准只有一个——基因在后代中留下副本的数量。一个对物种长期发展毫无益处的特征,只要它能在当下环境中帮助个体多生育后代,就会被自然选择保留。雄孔雀那条华丽但累赘的尾羽就是典型的例子:它对孔雀的日常行动是累赘,却能在求偶竞争中赢得优势,于是被保留了下来。而一旦环境改变,比如出现专门追踪鲜艳颜色的新天敌,这个“优势”可能立刻变成致命的负担。
适应性在进化生物学中的定义,是个体将自己的基因传递到后代的能力,而非通常意义上的“强壮”或“聪明”。一个身体强壮却不生育的人,进化适应性为零;一个体弱多病但子孙满堂的人,进化适应性极高。
利他行为的进化解释
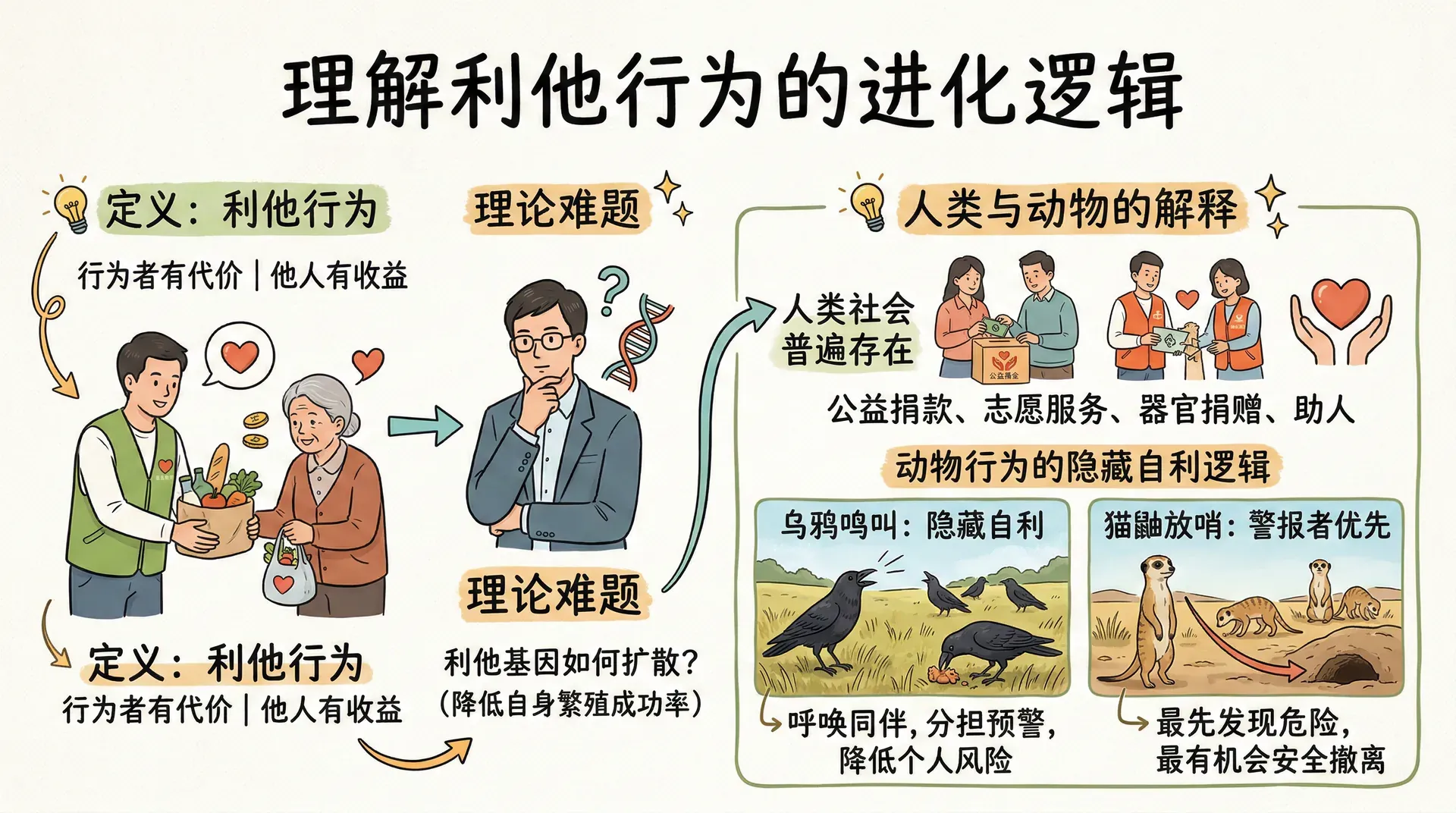
利他行为是指对行为者本人有代价、但对他人有益的行为。从进化角度来看,这类行为面临一个理论上的难题:如果一个基因使个体倾向于帮助他人,却在这个过程中降低了自身的繁殖成功率,那么这个基因如何能在种群中扩散?
在人类社会中,利他行为相当普遍。公益捐款、志愿服务、器官捐赠,乃至在考试前向竞争者讲解题目,这些都是以一定代价换取他人利益的行为。在其他动物中,表面上的“利他”行为往往可以找到隐藏的自利逻辑。乌鸦发现食物时会鸣叫召集同伴——这看起来是在“分享”,但地面上的单独觅食者面对猫和其他天敌时极为脆弱,呼唤同伴其实是在为自己争取更多的预警眼睛。猫鼬群体中确实有个体站岗放哨,但最先发现危险并发出警报的那只,往往也是最有机会安全撤离的那只。
真正意义上的利他行为——承担真实风险或代价、却不期待任何回报——在非人类动物中极为罕见,这本身就是一个值得深思的进化现象。
亲缘选择与互惠利他
理解利他行为如何在进化中存续,有两个关键机制:
-
亲缘选择。如果一个基因使个体倾向于保护自己的亲属,而亲属又携带与自己相同的基因——包括这个利他基因本身——那么这个基因就可以通过亲属的繁殖来传播,即便帮助者自身付出了代价。亲缘程度越近,共享基因的比例越高,帮助行为的进化收益也就越大。这解释了为什么父母对子女的牺牲在动物界如此普遍,也部分解释了为什么人们通常对直系亲属比对陌生人更愿意伸出援手。
-
互惠利他主义。即“你帮我,我帮你”的合作逻辑。但研究发现,人类的互惠利他并不局限于直接回报。人们不仅愿意帮助曾经帮助过自己的人,也愿意帮助那些被观察到曾经帮助过他人的人——也就是说,声誉本身就足以触发合作意愿。这种机制在人口规模较小、成员长期互动的社群中效果最为显著,因为声誉的积累需要人们能够识别彼此并记住各自的行为记录。
这也从侧面解释了为何利他行为在人类中比在大多数其他物种中更为普遍:人类拥有长期记忆、语言沟通和复杂的社会网络,这些能力使得声誉系统得以有效运作,互惠合作的范围也因此可以扩展到直接亲属之外。
亲缘选择和互惠利他并不是对立的机制,而是在不同情境下各自发挥作用的两条路径。前者依赖于基因的共享,后者依赖于社会记忆与声誉。两者共同构成了人类合作行为的进化基础。
动物研究的伦理争议
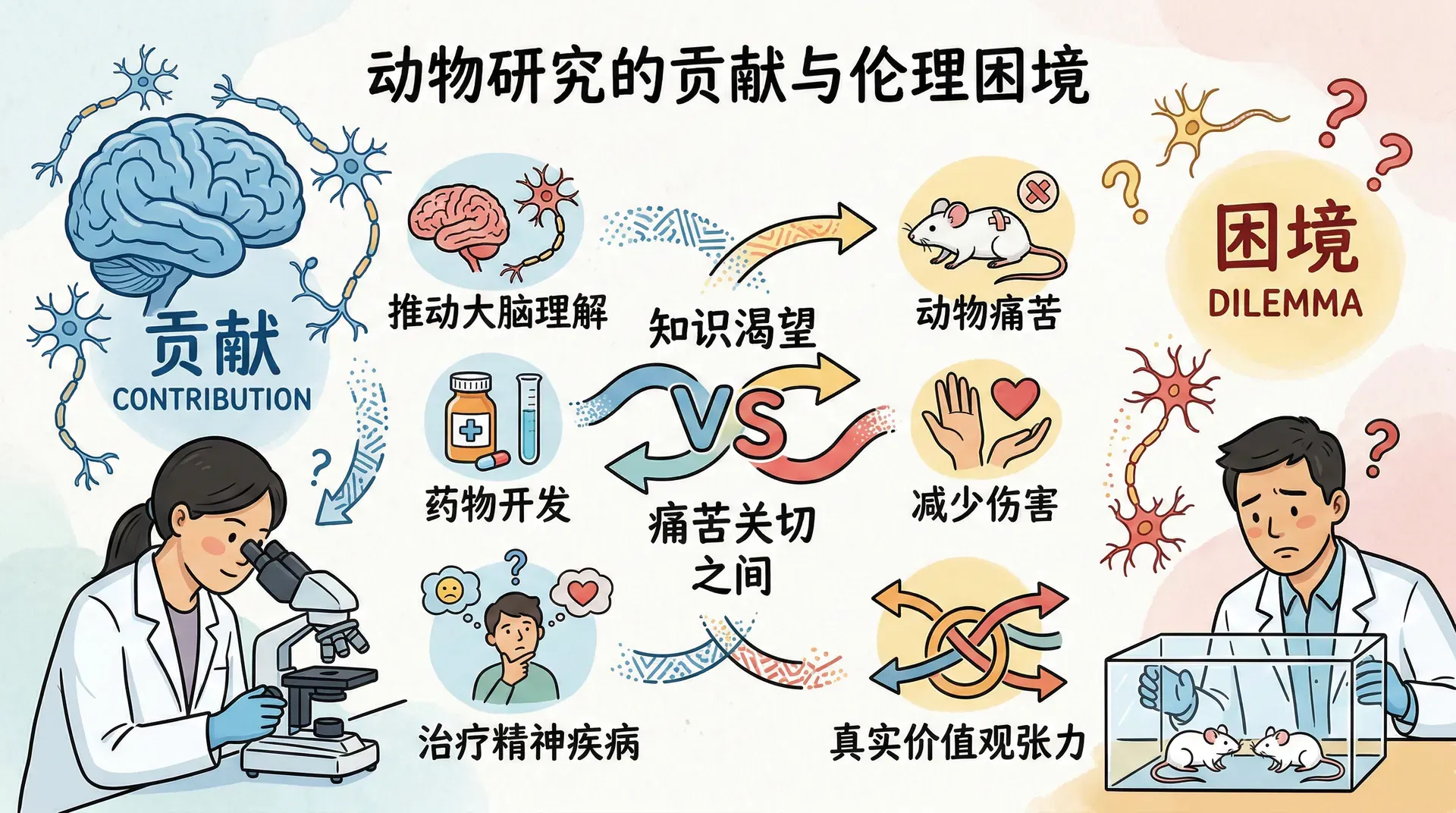
实验室动物研究为我们理解大脑和行为提供了大量不可或缺的知识。从脊髓反射的发现到神经递质功能的阐明,再到各类精神疾病治疗药物的开发,背后几乎都有动物实验的贡献。然而,这种研究同时也带来了一个真实的伦理困境:一方面是对知识的渴望,另一方面是对减少动物痛苦的关切,两者之间并不总是能够和平共处。这不是非黑即白的问题,而是两种真实价值观之间的张力。
研究非人类动物的理由
主要从事人类行为和大脑研究的生物心理学家,为什么会大量使用动物作为研究对象?这背后有几个务实的原因。
行为的基本神经机制在脊椎动物之间高度保守,许多在小鼠或大鼠身上发现的原理,同样适用于人类。较简单的神经系统往往更容易操作和分析,就像工程师在研究复杂设备时,有时更愿意先从结构相似但功能更简单的原型入手。
动物本身也具有独立的研究价值。蝙蝠如何在完全黑暗中用回声定位精确追踪猎物?候鸟迁徙时如何凭借内部导航系统跨越数千公里?这些问题不仅有助于理解动物行为本身,也为认识神经系统的普遍工作原理提供了独特视角。
此外,对动物的研究为重建人类进化历程提供了比较依据。我们与黑猩猩共享约98%的基因,但在认知能力和行为上差异巨大,这种差异究竟源自哪些遗传变化,需要通过比较研究才能揭示。
还有一个现实的原因:某些实验在技术上或伦理上无法在人类身上进行。将微电极直接插入脑细胞记录神经放电、通过手术制造特定脑区损伤来观察行为变化、长期控制饮食和睡眠条件——这些操作在动物研究中是常规手段,但在人类中受到严格的法律和伦理限制。
非人类脊椎动物的大脑在化学成分和解剖结构上与人类高度相似,甚至一些无脊椎动物的神经元也遵循与人类神经元相同的基本电生理原理。对乌贼巨大轴突的研究,正是因为其直径远大于人类神经纤维,才使科学家得以首次记录到动作电位的完整波形。
不同立场的分歧所在
对动物研究的反对意见存在不同的程度。持“最小主义”立场的人认为,动物研究在某些条件下是可接受的,但应当受到严格约束——包括限制使用动物的数量、尽可能以计算机模拟或体外实验代替活体动物,以及持续改进实验程序以减少痛苦。这一立场在科学界内部也获得广泛认同,并已体现在各国的监管框架中。
在中国,凡是接受政府科研经费资助的机构,均须成立由兽医、科学家和社会代表共同组成的动物福利委员会,负责审查实验方案并监督执行。“减少、替代、改善”三原则是国际通行的动物实验伦理准则。
持“废除主义”立场的人则认为,任何以动物为代价的研究都不可接受,无论目的是否出于医学知识的增进。这一立场的核心主张是:动物拥有与人类同等的权利,既然人类受试者的参与以知情同意为前提,那么无法表达同意的动物就不应当被用于任何实验。
这两种立场之间的分歧,本质上是两种伦理原则的冲突:“避免伤害无辜生命”与“允许有限的伤害以换取更大的益处”。这不是一方正确、另一方错误的对立,而是两种各有内在逻辑的价值体系在碰撞。
伦理争议本身并不可怕,可怕的是讨论被极端化,以至于持中间立场的研究者不得不沉默。许多从事动物研究的科学家对动物福利抱有真实的关切,但在高度对立的舆论氛围中,表达这种关切反而可能被误读为立场软化。
总结
生物心理学的核心假设是:大脑的活动状态就是心理状态本身。当你看到一朵花、感到一阵恐惧、做出一个决定,这些体验都对应着神经系统中某种特定的活动模式。这不是说“大脑控制了你”,而是说大脑与你是同一件事,只是从不同的描述层次来观察。
这一假设将我们引向一系列具体的研究问题:哪些脑区的损伤会导致哪些特定的行为变化?特定的激素或神经递质水平如何影响情绪和认知?遗传差异如何预测个体在心理特征上的差异?早年经历如何通过表观遗传机制改变大脑的长期状态?
目前可以确定的是,基因对行为的影响既不是决定性的,也不是可以忽略的。某些能力,如对特定化学物质的味觉敏感度,受单一基因的影响非常明显;更多的行为特征,如性格倾向、认知风格、精神疾病的易感性,则是多个基因与多种环境因素共同作用的产物。进化为人类提供了一套基本的行为倾向,但这套倾向本身就包含了高度的灵活性——我们能够在不同的文化和社会规范中调整自己的行为,这本身就是进化的结果。
研究人类行为的遗传和进化基础面临诸多困难。研究者无法控制人类的繁殖配对,无法像动物实验那样系统地操控基因或环境变量,对远古祖先生活条件的了解也十分有限。这些限制意味着,在这个领域,谨慎的结论比自信的断言更接近真相。
理解行为的生物学基础,并不是要证明人的命运由基因决定,而是要为我们认识自身、改善心理健康、设计有效干预手段提供更坚实的科学依据。知道某种倾向来自何处,是改变它的第一步。