感官信息处理与大脑认知
凝视齐白石的《虾》时,那水墨晕染如何在大脑中化作生动形象?聆听《高山流水》的古琴声时,那些音符又是如何触动心弦?炎热夏日品尝一口清甜西瓜,那份清爽又是如何传递到感官世界的?这些日常体验背后,隐藏着大脑复杂精妙的信息处理机制。大脑如同一位身处暗室的指挥家,却能通过各种感官信号把握外界的精彩世界。
感知的本质与功能
感知是理解和把握外部世界各种物体和事件的整体过程,涵盖感受、理解、识别刺激并为行为反应做准备。感知并非单纯被动地接收信息,而是一个主动建构的过程——大脑不断将零散的感官输入整合为有意义的完整体验。
感知过程承担着两大使命:在危险来临时及时警觉,以及引导人趋向愉悦体验。在北京胡同漫步时,突然响起的自行车铃声会让感知系统立即启动警觉反应;而路边烤鸭店飘来的香味,又会引导人去寻找这份美味。这两种截然不同的反应,都来源于同一套感知系统,只是触发了不同的处理机制。
感知与感觉是两个不同的概念。感觉是感官对物理刺激的直接反应,是原始的神经信号;感知则是大脑对这些信号的解读与组织,赋予其意义。听到一段旋律是感觉,认出那是《茉莉花》则是感知。二者相辅相成,共同构成人类认识世界的基础。
感知的三个核心阶段
感知并非一蹴而就,而是经历三个相互衔接的阶段,每个阶段都在前一阶段的基础上对信息进行更深层次的加工。理解这三个阶段,是掌握整个感知心理学的关键起点。
第一阶段是感觉阶段。 环境刺激作用于感觉受体时,受体将物理能量转换为神经冲动。光线进入眼睛、声音传入耳朵、气味分子接触鼻腔,感觉细胞收集这些原始信息并传递给大脑。这一阶段提供的是最基本的感官数据,尚无任何意义的附加。
第二阶段是感知组织阶段。 大脑将感官信息与已有知识结合,形成对外部刺激的内在表征。这个过程帮助人估计物体的大小、形状、运动、距离和方向。欣赏中国山水画时,大脑会自动将墨点和线条组织成连绵山峦与流动水势的立体图像,而不是一堆没有意义的色块。
第三阶段是识别认知阶段。 这一阶段赋予感知以意义,完成从“物体看起来是什么样”到“这是什么”和“有什么功能”的转变。看到圆形物体时,识别过程判断它究竟是篮球、硬币、时钟还是月亮;看到一张面孔时,识别过程确定这个人是陌生人还是认识的朋友。
三个阶段虽然在描述上是分开的,但在大脑中几乎是同时运作的。感觉、组织和识别彼此交织,高层次的认知过程可以反向影响低层次的感觉加工,这正是感知系统精妙之处。
感觉认知的基础原理
准确的感知是生存的基础——寻找食物、躲避危险、与他人交往,无不依赖对世界的可靠感知。心理学家为了揭示这些感知背后的规律,发展出一套系统的研究框架,从物理刺激到心理体验,逐步建立起感觉认知的基础理论。
远端刺激与近端刺激
坐在客厅环顾四周时,物体反射的光线在视网膜上形成图像。视网膜图像与真实环境存在根本差异:视网膜图像是二维的,真实环境是三维的。这种差异带来了一个有趣的困惑——房间里的桌子、地毯、窗户和画框在现实中都是矩形,但只有窗户在视网膜图像中呈现为矩形;画框是梯形、桌面是不规则四边形、地毯则被分成若干区域,边数远超实际。
心理学家将物理世界的真实物体称为远端刺激,将视网膜上的光学图像称为近端刺激。感知的主要任务,就是从近端刺激信息中还原出远端刺激的真实特征。
以挂在墙上的画框为例:感觉阶段对应视网膜图像中的二维梯形;感知组织阶段将这个梯形理解为三维空间中远离观看者的矩形;识别阶段才最终将这个矩形物体认定为一幅画作。这三个阶段的连续推进,体现了大脑从混乱的近端刺激中重建真实世界的能力。
心理物理学与感觉阈限
心理物理学研究的是物理刺激与心理体验之间的关系。工厂里的火警需要多响亮才能在机器声中被工人听到?茶叶中加多少茉莉花才能让饮茶者察觉到香气的变化?德国物理学家费希纳创立了这一学科,建立了物理刺激强度与感觉体验强度之间的关联体系。
绝对阈限是其中最基本的概念,指产生感觉体验所需的最小物理能量。研究者会让人在黑暗中察看微弱光线、在安静环境中辨听轻微声音,通过测量不同强度下的觉察情况来确定阈限值。人类各感官系统的绝对阈限差异极大,下表列出了各感官在理想条件下的检测能力:
感觉系统对环境变化比稳定状态更为敏感。感觉适应是指受体细胞在持续刺激下逐渐失去反应能力的现象——在阳光下待久了不再感到刺眼,走进飘有气味的房间后逐渐感觉不到异味,都是感觉适应在生活中的体现。这种机制帮助感知系统将注意力集中在新出现的变化上,而非消耗在不变的背景中。
信号检测与韦伯定律
阈限的测量并非单纯的感觉问题,还受到决策倾向的影响。在光线检测实验中,如果正确报告“有光”可以获得奖励,而错误报告则受到惩罚,这种奖惩结构会直接影响人的反应方式。信号检测理论将感觉过程与决策过程区分开来:前者决定感官的敏感性,后者决定在不确定情况下倾向于报告“有”还是“无”。
差别阈限是另一个重要概念,指能够被感知为差异的两个刺激之间的最小物理差异。韦伯在1834年发现了一条规律:差别阈限并非固定值,而是与标准刺激强度成固定比例。刺激越强,需要更大的物理增量才能让人感觉到变化。
提着25公斤的行李时,多加0.5公斤才能察觉到重量增加;提着50公斤时,则需要多加1公斤才有同样的感觉。增量不同,但比例始终保持一致,约为0.02。这条规律在饮料配方、音响设备乃至工程设计中都有广泛的应用价值。
能量转换机制
感觉的核心奥秘在于物理能量如何引发特定的心理体验——光的波长如何让人体验到颜色?声音的振动如何让人感受到音调的高低?这一转换过程被称为换能,即将一种物理能量转变为神经冲动的过程。
由于所有感觉信息最终都转换为相同类型的神经冲动,大脑并非通过冲动本身的性质来区分感觉,而是通过激活特定的皮层区域来产生不同的感觉体验。视觉皮层受到刺激产生视觉,听觉皮层受到刺激产生听觉——感觉的质量取决于信号抵达的目的地,而非信号的形式本身。
视觉系统

视觉是人类最复杂、最依赖的感觉通道,占据了大脑皮层约30%的处理资源。良好的视觉能力帮助人发现远处的目标或危险,察觉环境的细微变化并及时调整行为。从简单的光线进入眼球,到大脑中形成完整的视觉世界,这一过程涉及从眼球结构到神经通路的一系列精密机制。
眼睛的结构与成像
眼睛是视觉系统的感光器官,其结构与照相机高度相似。光线依次通过角膜、前房液体、瞳孔进入眼内。虹膜通过调节瞳孔大小来控制进光量——光线强时瞳孔收缩,光线弱时瞳孔放大。豆形的晶状体通过改变自身厚度来实现聚焦:变薄时聚焦远处物体,变厚时聚焦近处物体。光线最终汇聚在覆盖眼球后壁的视网膜上,成像过程就此完成。
近视和远视是最常见的调节问题。近视者的晶状体聚焦点落在视网膜前方,无法看清远处;远视者则相反。随着年龄增长,晶状体逐渐失去弹性,45岁以后清晰聚焦的近点开始逐渐后退,这就是中老年人需要使用老花镜的原因。
视网膜的感光机制
视网膜是眼球内真正完成光电转换的结构,由视杆细胞和视锥细胞两类感光受体构成。视网膜拥有约1.2亿个视杆细胞,分布于视网膜周边区域,专门在暗光环境下工作,对光线极为敏感,但无法分辨颜色。约700万个视锥细胞则集中分布于视网膜中央的中央凹区域,负责在明亮光线下处理细节和色彩信息。
每次关灯之后,从看不清周围环境到逐渐能够分辨物体轮廓,这一过程就是视杆细胞的暗适应过程,通常需要20至30分钟才能完全完成。中央凹是视觉最敏锐的区域,正是因为该处视锥细胞密度最高,所以阅读文字时总是要将目光直接对准文字,而非用余光扫视。
视神经离开眼球的区域没有任何感光细胞,形成所谓的盲点。但由于两眼的盲点位置不同,且大脑会主动填补缺失的信息,日常生活中几乎察觉不到盲点的存在。
视觉信息的皮层处理
从视网膜出发的视神经信号,经由丘脑的外侧膝状体中继,最终传送到枕叶的初级视觉皮层。视神经在视交叉处交叉分束,使得每侧大脑都能接收到来自双眼的信息。初级视觉皮层接收信号后,会将其分流至两条平行的处理通路:一条负责识别物体的形状和颜色,被称为“是什么”通路;另一条负责定位物体在空间中的位置,被称为“在哪里”通路。
视觉系统由多个独立的子系统组成,分析同一视网膜图像的不同方面。休伯尔和维瑟尔因发现视觉皮层细胞的感受野特性而获得1981年诺贝尔生理学或医学奖。他们发现,皮层中的简单细胞对特定方向的光条反应最强,复杂细胞要求光条移动才能激活,超复杂细胞则对特定长度或角度的移动光条做出反应。视觉皮层的这种层级化组织,正是大脑从简单线条中识别出复杂形状的神经基础。
色彩感知与颜色视觉理论
颜色体验的物理基础是光的波长。可见光是电磁频谱中波长约为400至700纳米的一段,波长不同,产生的颜色体验也不同——短波长呈现紫蓝色,长波长呈现红橙色。颜色并不存在于物体本身或光线之中,而是存在于感觉系统对波长的解释中。红色中国结之所以看起来是红色,是因为它反射了波长约为700纳米的光线,而大脑将这一波长的光解读为红色。
所有颜色体验可以用三个维度来描述:色调(由波长决定的颜色种类,如红、绿、蓝)、饱和度(颜色的纯净程度,灰色接近零饱和度)、明度(颜色的亮暗程度)。人类能够区分约700万种不同的颜色,但大多数人仅能说出其中极少数的名称。
关于颜色视觉的机制,历史上存在两种重要理论。杨-亥姆霍兹三色理论认为人眼有三种颜色受体,分别对红、绿、蓝三种波长敏感,其他所有颜色都是这三种基色不同比例的组合。然而,这一理论难以解释色彩适应后出现的互补色残像,以及色盲者总是成对无法区分红绿或蓝黄这两类颜色的现象。
赫林的对立过程理论则认为,所有颜色体验来自三个互相对立的系统:红对绿、蓝对黄、黑对白。
现代研究表明,两种理论并不矛盾,而是描述了视觉系统不同层级的处理机制。在视网膜层面,三种类型的视锥细胞分别对不同波长的光最为敏感,这与三色理论一致;而视网膜的神经节细胞则按照对立过程整合三种视锥的输出——一些细胞受红色光兴奋、受绿色光抑制,另一些细胞恰好相反。两种机制在不同阶段共同参与,才构成了完整的色彩感知系统。
听觉系统

听觉与视觉的功能高度互补。声音往往先于视觉信号到达——身后传来的脚步声、门后的对话声,都是视觉无法捕捉而听觉能够先行察觉的信息。听觉系统不仅负责接收语言和音乐,更承担着在三维空间中定位声源、协助视觉系统校准注意方向的重要功能。
声音的物理特性与心理维度
拍手、吹口哨、敲击桌面——这些动作之所以能产生声音,是因为物体的振动扰动了周围的空气分子,产生压力变化,以每秒约340米的速度向四周传播,形成疏密相间的声波。真空中没有可以被推动的分子,因此声音无法在真空中传播。
声波的频率和振幅是决定声音特性的两个核心物理参数,它们分别对应三个心理维度中的音调和响度:
-
音调由频率决定,高频声波产生高音调,低频声波产生低音调。人类能够感知的频率范围约为20至20000赫兹,钢琴88个琴键涵盖的频率仅在30至4000赫兹之间。频率与音调的关系并非线性——在低频段,仅需增加几赫兹就能感受到明显的音调变化;在高频段,则需要大得多的增幅才能产生同样的感觉变化。
-
响度由振幅决定,振幅越大声音越响亮,振幅越小则越轻柔。响度以分贝为单位来衡量。生活中常见声音的分贝水平如下表所示:
-
音色反映声波的复杂组成,是区分同一音调下钢琴与二胡声音的依据。自然界中大多数声音都是多个频率叠加的复杂波,而非单一频率的纯音。
长期暴露在90分贝以上的环境中,会对听觉系统造成不可逆的损伤。建筑工地、纺织车间的工人以及长期使用耳机大音量听音乐的人群,都面临较高的职业性或习惯性听力损失风险。
声音转变为神经信号的过程
声音从进入耳朵到被大脑处理,需要经历一系列精密的能量转换过程。
振动的空气分子沿外耳道传播,到达鼓膜后引起鼓膜的相应振动。外耳廓的形状有助于收集和引导声波,使之集中进入耳道。
鼓膜的振动依次传递给中耳的三块听小骨——锤骨、砧骨和镫骨。这三块人体中最小的骨骼构成一条机械传导链,将振动信号放大并集中传递至内耳的卵圆窗。
镫骨推动卵圆窗,使耳蜗内的淋巴液产生波动。耳蜗是一段充满液体的螺旋形管道,其中的基底膜在液体波的推动下产生波状运动,刺激附着其上的毛细胞发生弯曲。毛细胞是听觉系统真正的感受器,其弯曲触发神经电信号的产生。
毛细胞产生的神经冲动经由听神经传入脑干,再经过一系列中继核团的处理,最终抵达颞叶的听觉皮层,在那里完成对声音的高级分析与识别。与视觉系统类似,来自一侧耳朵的信号也会传送至大脑的双侧,从而增强声音定位的精度。
音调感知的位置理论与频率理论
听觉系统如何将不同频率的声波转化为对应的音调感觉?研究者提出了两种互补的理论来解释这一问题。
位置理论由亥姆霍兹提出,后经贝克西实验验证,贝克西因此获得1961年诺贝尔奖。该理论认为,音调的感知取决于基底膜上振动最剧烈的位置——高频声音使基底膜靠近卵圆窗的底部区域振动最大,低频声音则主要激活基底膜远端的顶部区域。不同位置的毛细胞激活,产生不同音调的感觉。
频率理论则认为,基底膜的整体振动频率直接编码了声音的频率信息——100赫兹的声波使基底膜每秒振动100次,神经元以相同的频率放电,大脑由此识别音调。然而,单个神经元的放电速率存在上限,无法超过每秒1000次,因此频率理论对高频声音的解释存在困难。为此,研究者提出了齐射原理——多个神经元交替放电,共同编码2000赫兹乃至更高频率的信号。
两种理论实际上描述了同一系统在不同频率范围内的工作方式。频率理论较好地解释了5000赫兹以下的低中频声音的编码方式;位置理论则更好地说明了1000赫兹以上高频声音的感知机制。二者相互补充,共同构成完整的音调感知理论框架——这与颜色视觉中三色理论和对立过程理论的关系如出一辙。
声音的空间定位
穿越上海人民广场时,远处有人呼唤名字,人们往往能够准确判断声音来自哪个方向,并迅速转身。这种声音定位能力依赖两种协同工作的机制。
第一种机制是双耳时间差。来自右侧的声音会比到达左耳早几百微秒就先抵达右耳。听觉系统中存在专门比较双耳到达时间差异的神经元,大脑根据这一时间差对声源的方位做出精确推算。
第二种机制是双耳强度差。头颅本身会对声音产生遮蔽效应——声源同侧耳朵接收到的声音强度略大于对侧。神经系统中有专门检测双耳强度差异的细胞,与时间差机制协同工作,共同实现对声源位置的精准定位。
其他感觉系统

视觉和听觉虽然是研究最为深入的两个感觉通道,但人类完整的感觉系统远不止这两种。嗅觉、味觉、皮肤感觉、疼痛以及感知身体状态的内部感觉,共同构成了感知世界的完整图景。这些感觉彼此配合,在不同情境下各司其职,缺一不可。
嗅觉
气味分子进入鼻腔后,与嗅觉纤毛膜上的受体蛋白结合,引发神经冲动。仅仅8个气味分子就能触发一次神经冲动,但至少需要刺激到40个神经末梢,才能产生可以被意识到的气味感觉。这些冲动经由嗅球传递到大脑的边缘系统和额叶区域,这也是为什么气味往往能够强烈唤起情绪记忆——闻到某种老家食物的香气,儿时的记忆便会扑面而来。
嗅觉系统具有一项独特的再生能力:无论是嗅觉受体细胞还是嗅球中的神经细胞,都能在一定程度上更新替代。这使得部分在脑损伤或感染后失去嗅觉的患者,有机会逐渐恢复嗅觉功能,这在其他神经感觉系统中是非常罕见的。
嗅觉在人类社会生活中的功能远不止于品味食物。许多文化习俗、节日氛围与特定气味密不可分——春节的鞭炮硫磺味、中秋月饼的桂花香、清明时节的青草气息,都通过嗅觉与深层的文化记忆相连。
味觉
味觉与嗅觉的关系极为密切,二者协同工作,共同构成人对食物的完整感受。感冒鼻塞时感觉食物“没有味道”,正是因为嗅觉通道受阻,味觉信号失去了嗅觉的配合。实际上,美食家对食物的精细评判,相当一部分来自嗅觉而非味觉本身。
舌头表面分布着数千个味蕾,每个味蕾中包含约50至150个味觉受体细胞。研究表明,味觉受体细胞能够识别五种基本味觉质量:甜、酸、苦、咸和鲜味。其中鲜味是较晚被确认的一种,对应谷氨酸钠的味道,自然存在于肉类、海鲜、豆腐和陈年奶酪中,也是中餐调味的重要基础之一。五种基本味觉并非彼此孤立,而是以不同的比例组合,形成人类对食物风味的丰富感知。
皮肤感觉与疼痛
皮肤是人体面积最大的感觉器官,承担着保护机体、调节体温和传递触觉信息的多重功能。皮肤内分布着多种不同类型的感觉受体,分别对压力、振动、温度和痛觉做出反应。触觉小体在物体摩擦皮肤时最为活跃,触觉盘则对持续的稳定压力最为敏感。
疼痛是感觉系统中一种特殊而重要的存在,它是机体对有害刺激的警告信号。天生缺乏痛觉的人虽然免受疼痛之苦,却往往因无法感知伤害而使身体遭受严重损伤——烫伤、骨折、关节磨损,种种本可通过疼痛预警而避免的伤害,在他们身上频繁发生。
大脑中的内啡肽系统在疼痛调节中扮演着核心角色。内啡肽是一类由大脑自行合成的神经肽,与吗啡结合在同一受体位点上,具有天然的镇痛效果。针灸能够缓解疼痛,研究者认为这在相当程度上与针刺刺激引发内啡肽释放有关。安慰剂同样能够产生真实的镇痛效果,其机制也与内啡肽的释放密不可分。
关于疼痛如何在神经系统中被调控,门控制理论提供了一个重要的解释框架。该理论认为,脊髓中存在一种神经“门”机制,能够根据来自大脑的信号决定是否放行疼痛信号。这解释了为何情绪状态、注意力和心理预期能够显著影响疼痛的感受强度——受伤时心情平静往往比惊慌失措感觉更少的疼痛。
前庭觉与运动觉
前庭觉和运动觉是两种没有专属感觉器官的内部感觉,却对维持身体协调和空间定向不可或缺。
前庭觉告知大脑头部相对于重力的朝向和加速度变化。其感受器位于内耳的半规管和耳石器中——充满液体的管道在头部转动时产生液体流动,压迫其中的毛细胞,由此产生方向感和平衡感。乘坐电梯或高速列车急刹时产生的眩晕感,正是前庭系统对加速度变化的反应。
运动觉则通过分布在关节、肌肉和肌腱中的感受器,持续向大脑报告身体各部分的位置和运动状态。闭上眼睛仍然知道手臂举起还是放下,知道腿在弯曲还是伸直,这种对身体状态的感知能力完全依赖运动觉。没有运动觉的协助,大脑发出的每一个运动指令都将无法得到有效反馈,精细动作的完成将变得极度困难。
知觉组织

感觉受体每时每刻都在向大脑输送海量的原始信号——仅视网膜就有一亿多个感光细胞同时工作。若没有一套将这些信号组织为连贯整体的机制,外部世界在感知中将只是混乱的光影片段。知觉组织正是大脑将原始感觉信息重新整合为有意义的感知场景的过程,是从感觉到感知的关键一环。
选择性注意
在任何一个时刻,感觉系统都在同时接收来自环境的大量信息——墙上的裂缝、窗外的车声、空气中的气味、桌面的纹路。然而意识的容量是有限的,无法将所有这些信息同等对待。注意的功能就是在全部可用信息中进行筛选,将意识资源集中到当前最相关的对象上。
注意力的分配受两类机制控制。目标导向的注意是主动性的,反映人根据自身目标和意图所做的选择——在书店里专注寻找某一类书目时,目光会自动在书架上扫描相关字样,其他信息则被过滤掉。刺激驱动的注意则是被动性的,由环境中某些具有突出特征的刺激自动捕获——正在专心阅读时,窗外突然传来一声巨响,注意力会不由自主地被拉走。
人脸是最强效的注意捕获刺激之一。研究发现,即便在完成某项具体任务时,如果任务中出现人脸,即使忽略人脸能获得更好的成绩,注意力依然会被人脸吸引。这一现象表明,大脑对具有进化意义的生物性刺激保持着自动化的优先处理通道。
格式塔知觉组织原理
20世纪初,德国格式塔心理学家系统研究了感觉信息如何被组织为完整的知觉整体。他们的核心主张是:心理现象只有作为有组织的整体才能被正确理解,整体不等于部分之和,而往往远大于部分之和。
格式塔心理学家通过改变视觉刺激阵列中的单一因素,总结出若干普遍适用的知觉组织原则:
这些原则并非相互独立,而是同时作用于感知过程。现实生活中的知觉场景往往需要多种原则共同参与,才能形成清晰、稳定的知觉组织。
运动知觉与深度感知
感知的任务不仅是识别静态的物体,更要追踪运动中的目标,以及理解三维空间中物体之间的深度关系。
运动知觉依赖于对视网膜图像随时间变化的分析。当一个物体在视野中移动时,其在视网膜上的图像位置也随之位移,大脑据此判断物体的运动方向和速度。走向对面的朋友时,对方在视网膜上的图像逐渐扩大,扩大的速率给出接近速度的直接信息。费现象揭示了大脑对运动的主动建构——当两个静止光点交替闪烁时,大脑会自动将其解读为一个光点的往复运动,而非两个分别闪烁的光点。
深度感知则依赖于多种来源的深度线索:
-
双眼线索来自两眼视角的差异。由于两眼相距约6厘米,它们接收到的是略有不同的视角。这种双眼视差随物体距离的增大而减小,大脑通过测量视差的大小来推算物体的远近。此外,眼球向内转动的聚散度也提供距离信息——注视极近的物体时,眼球必须大幅内转,大脑从眼部肌肉的张力中获得距离判断的依据。
-
单眼线索不依赖双眼视差,仅凭单眼也能感知深度。中国传统绘画中早已熟练运用这些原理:近大远小、遮挡关系、线条透视、纹理渐变、大气透视——远处的山峦颜色偏灰偏蓝,层叠的山脉依次后退,这些都是单眼深度线索在艺术中的自觉运用。
知觉恒常性
将手持的课本从正常阅读距离推远一倍,课本在视网膜上的图像面积会缩小为原来的四分之一,但人不会认为课本变小了。这种尽管视网膜图像发生变化、但感知仍保持稳定的现象,被称为知觉恒常性。
大小恒常性使人能够在物体远近变化时仍准确判断其真实大小,这依赖于大脑将视网膜图像大小和距离信息综合考虑的能力。形状恒常性使人能够识别各种角度和朝向下的同一物体——从侧面看到的碗口是椭圆形,但始终被识别为圆形的碗口。明度恒常性则使人能够在不同光线条件下稳定感知物体的明暗属性——白色宣纸在夜间反射的光量远少于灰色纸张在正午阳光下的反射量,但白纸始终被感知为比灰纸更白。
恒常性机制的本质是大脑的“折扣”能力:它能够从感觉输入中剔除环境因素(距离、角度、光线强度)的影响,还原出物体的本征属性。这一能力对于在复杂多变的环境中做出可靠判断至关重要。
错觉与知觉的局限
知觉恒常性机制通常运作得非常成功,但在某些精心构造的刺激条件下,大脑的推断会产生系统性偏差,导致错觉的产生。错觉与个人经历无关,同一文化背景的人面对同一刺激会产生相同的错觉,这说明错觉植根于感觉系统共同的神经机制,而非个体的想象。
缪勒-莱耶错觉是最经典的视觉错觉之一:两条等长线段,一条末端附有内敛的箭头,另一条末端附有外展的箭头,前者看起来明显比后者短。这一错觉来源于大脑将箭头末端解读为建筑物角落的经验习惯——内敛箭头类似楼角向外凸出,外展箭头类似墙角向内凹入,大脑的深度判断机制介入,使两条线段产生了长度差异的错觉。
错觉并不只是心理学实验室里的趣味现象,它在实际生活中具有广泛的应用。室内设计师和建筑师利用视觉错觉来改变空间感受——浅色的墙壁、纵向的条纹图案、适当的家具尺寸搭配,都能让小房间显得更宽敞。舞台美术和电影特效也大量运用知觉规律,制造出远比实际更宏大的视觉效果。
识别与认知过程
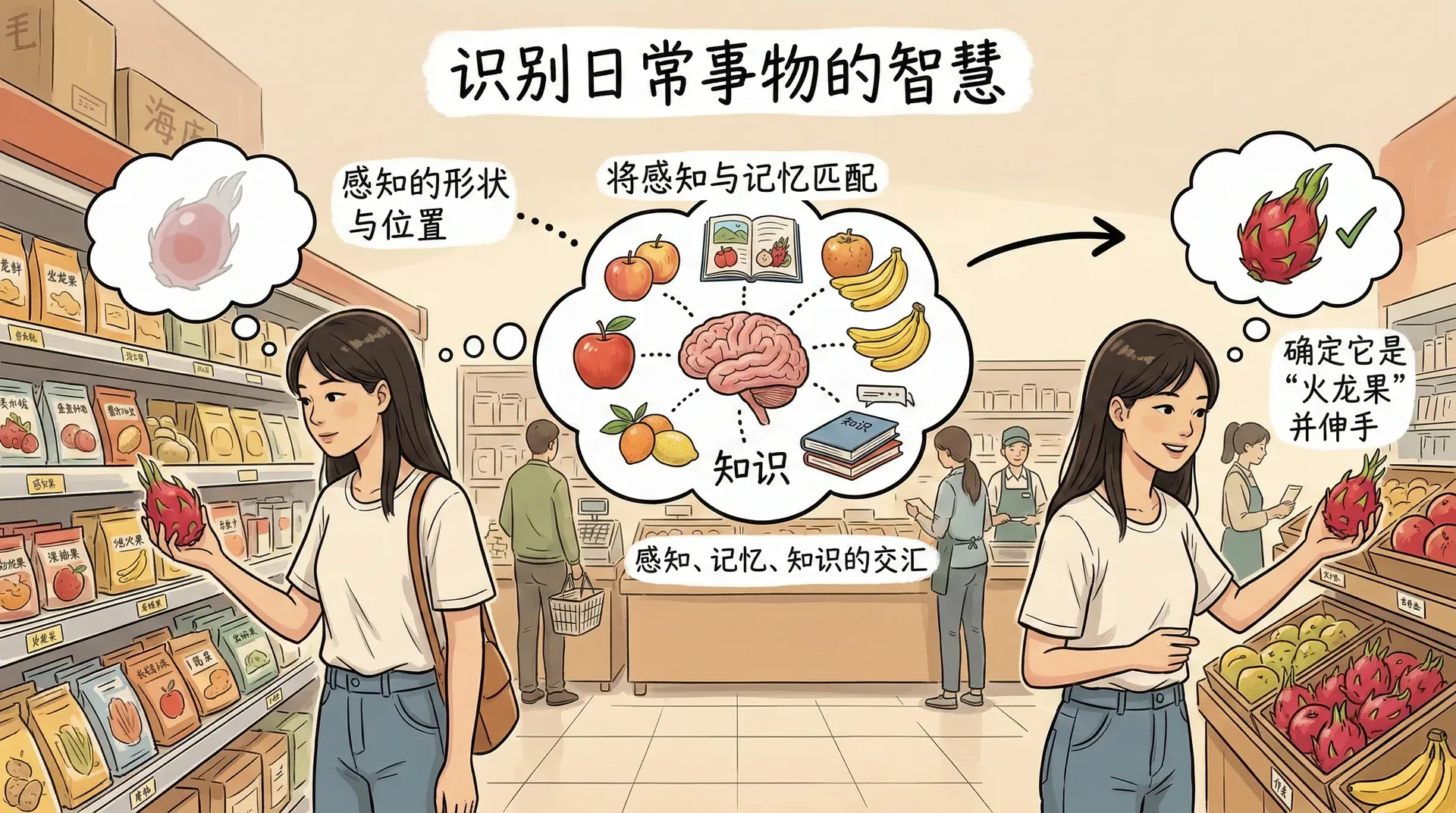
知觉组织过程完成之后,大脑已经知道视野中存在一个什么形状、处于什么位置的物体,但这还不足以指导行为。要对外界做出恰当反应,还需要识别——将当前感知到的内容与记忆中储存的知识匹配,从而确定“这是什么”。识别与认知是感知过程的最终阶段,也是感知与记忆、知识相互交汇的地方。
自下而上与自上而下的加工
识别过程可以从两个方向运作。自下而上加工是从感觉数据出发、逐级抽象的识别路径:感觉受体探测到物理信号,信号在神经系统中经过层层分析,最终被归类为某个具体的对象。这种加工不依赖先前的知识和期望,完全由输入数据驱动,因此也被称为数据驱动加工。
自上而下加工则是从已有的知识和期望出发,主动引导感知的过程。参观成都熊猫基地时,即便只看到一团黑白相间的毛茸茸物体,人们也会立即将其识别为大熊猫,而不会误以为是某种不知名的动物——背景知识在识别启动之前就已经缩小了可能性的范围。当期望和背景影响知觉时,这种加工方式就在发挥作用。
自上而下加工涉及过去经验、知识积累、当前动机和文化背景对感知的综合影响。它使得感知不仅仅是对外部信号的被动接收,而是一个主动的意义建构过程。正因为此,同一幅画在不同观者眼中可能引发截然不同的情感共鸣;同一段音乐在受过训练和未受过训练的听者耳中,感知到的细节和层次也大相径庭。
语音修复现象清楚地展示了自上而下加工的作用。在嘈杂环境中交谈时,语音信号经常被背景噪声遮盖,出现物理上的缺失,但听者几乎不会意识到这些空缺,大脑会根据语义和语境自动填补缺失的内容,使对话仍然流畅进行。
定势对识别的影响
感知的结果并不总是由感觉输入单方面决定的。背景信息和认知定势会显著影响同一刺激的识别结果。定势是一种临时性的准备状态,使感知系统倾向于以特定方式解读信息。
在一项经典示范中,先呈现一组动物名称——狐狸、猫头鹰、蛇、火鸡、天鹅,再出现一个两义词“鸽子”,此时多数人会将其理解为鸟类名称;而先呈现一组人名——晓东、建华、志强、明辉,再出现“鸽子”,人们则更容易将其理解为人名“戴维”的谐音。
定势的形成来自背景的激活效应。这一原理在日常生活中无处不在——在医院候诊时,对身体任何轻微异常的感知都会被放大;在运动场上观战时,同样的肢体碰撞感知到的程度远低于日常情境。空间背景、时间背景和情绪状态,共同决定了感知者以何种方式接收并解读眼前的信息。
社交疼痛与躯体疼痛的神经重叠
感知不仅处理来自外部物理世界的刺激,也处理来自社会关系的情感信号。情感受伤的说法往往被视为比喻,但神经科学的研究表明,社会性痛苦与身体疼痛在大脑层面激活的区域高度重叠。
一项研究招募了40名在六个月内经历过非自愿分手的参与者,在功能性磁共振成像扫描下同时测量身体疼痛和社交疼痛的神经反应。身体疼痛条件下,研究者对参与者前臂施加接近忍受极限的热刺激;社交疼痛条件下,参与者一边看着前任的照片,一边回忆被拒绝时的感受。
扫描结果显示,两种条件下激活的大脑区域存在显著重叠——处理身体疼痛的感觉皮层,在回忆分手经历时同样被激活。这一发现表明,“分手很痛”并非单纯的比喻,而是有着真实神经基础的生理体验。
感知的整合与平衡
从最初的光线照射视网膜到最终完成对世界的理解,感知是一个融合了感觉、组织、识别的多层次过程。每一层次都不是孤立运作的,而是与其他层次持续相互影响。
最终的感知体验不仅取决于感觉器官接收到的物理信号,还受到个人经历、情绪状态、文化背景和当前期望的深刻塑造。感知者在这个过程中同时扮演两个角色:一方面如同观察者,从纷繁的感觉证据中提取规律、做出推断;另一方面如同设计者,主动将零散的信息组织成连贯一致的整体,拒绝混乱,寻求秩序。
纯粹依赖自下而上加工的感知,会将人束缚于当下的物理刺激,无法从过往经验中获益;纯粹依赖自上而下加工的感知,则可能使人活在期望构建的幻象中,错过真实世界传来的重要信号。健康的感知系统在两者之间保持动态平衡——既尊重来自世界的原始信号,又善用积累的知识与经验,以最优的方式理解和回应所处的环境。