基因表达
在四川卧龙自然保护区,研究人员发现了一些极其罕见的白色大熊猫。这些“白熊猫”全身雪白,眼睛呈现粉红色,与我们熟悉的黑白相间的大熊猫形成鲜明对比。是什么因素导致了这种白化现象呢?
遗传性状的表现是由基因决定的,而白化现象通常是由于色素合成基因发生突变造成的。
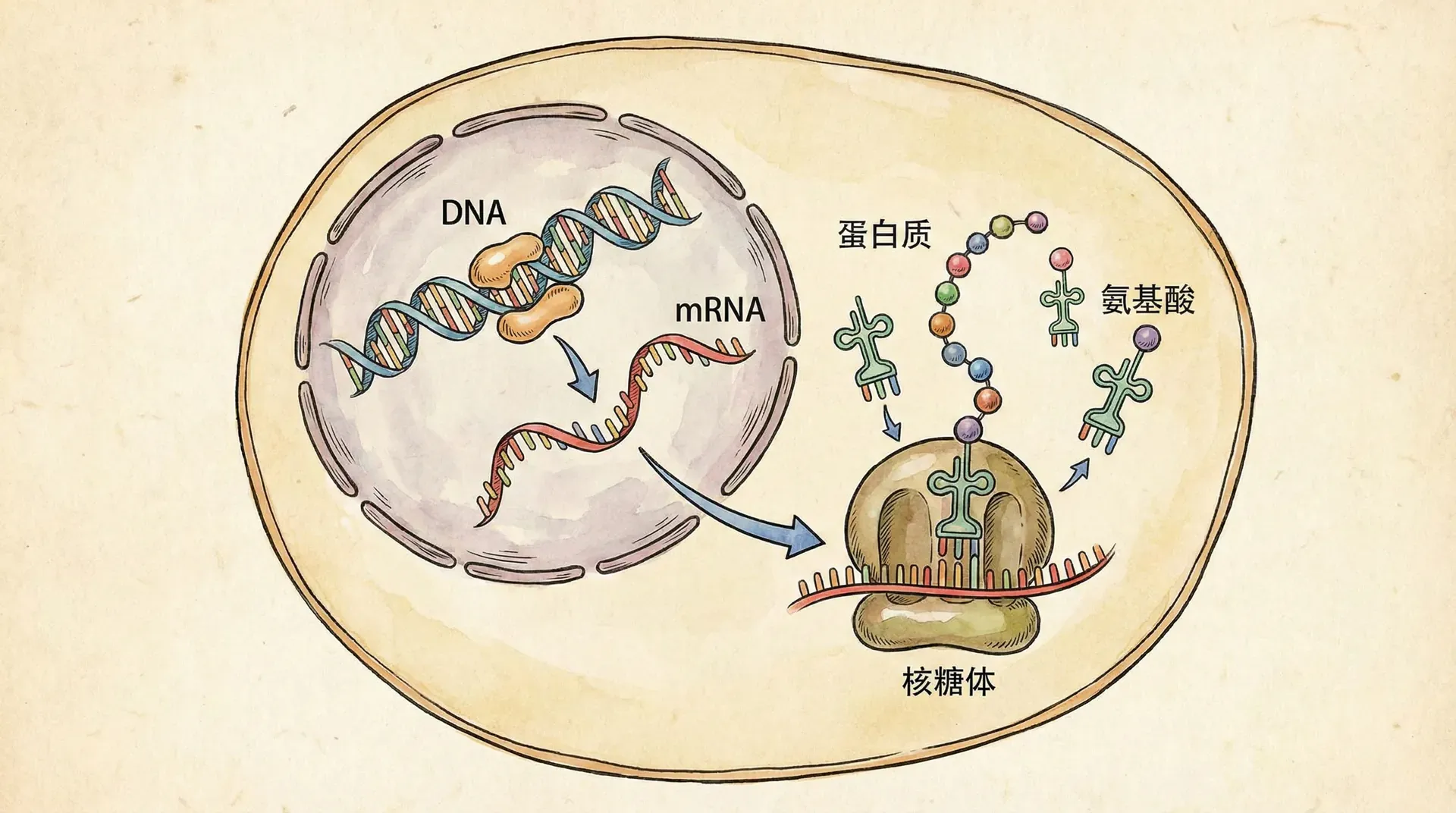
这个现象揭示了生物学中一个基本而深刻的原理:DNA中储存的遗传信息如何指导蛋白质的合成,进而决定生物的具体性状。基因表达就是指DNA通过指导蛋白质合成来实现遗传信息表达的过程。
在白化大熊猫的例子中,负责黑色素合成的酶蛋白出现功能缺陷,这种缺陷源于编码该酶的基因携带错误信息。从基因到蛋白质的信息传递包括两个主要环节:转录和翻译。
基因与蛋白质关系
早期的理论基础
20世纪初,英国医生加罗德通过对黑尿病这种遗传病的研究,首次提出了基因能够通过调控酶的产生进而影响性状表现的观点。他注意到黑尿病患者的尿液暴露于空气后会变黑,原因在于缺少分解特定化学物质(黑尿酸)的相关酶。加罗德据此推断:健康人可以正常合成该酶,而黑尿病患者出现这种现象,是因为其基因发生变异,导致缺乏这种酶。
酶假说的确立
几十年后,美国科学家比德尔与塔图姆选择面包霉菌作为实验模型,进一步验证了基因与酶间的联系。面包霉菌为单倍体生物,基因变异产生的表型效果能直接体现,非常适合做相关研究。
实验设计核心思路
面包霉菌通常可在含有无机盐、葡萄糖和生物素的基础培养基上独立合成生长所需所有氨基酸等营养。研究人员用X射线处理面包霉菌,诱导基因突变,再筛选出那些不能在基础培养基上存活的突变类型。
数据分析发现,各类突变株在补加特定物质后才能生长,从而揭示了精氨酸合成的各主要步骤分别由不同基因控制。
精氨酸生物合成途径中各基因作用的解析
上述结果有力地支持了“一基因一酶假说”,即每一个基因负责一种酶的形成。
假说的现代发展
进入分子生物学时代,科学家们对“一基因一酶”理论进行了补充和修正。当前较为普遍的观点是:
更精确地说,一个基因通常编码一条多肽链。许多蛋白质实际上由多条多肽链组成,而且每条多肽链往往由独立的基因编码。比如,血红蛋白中的两种多肽链正是由不同的基因分别编码的。
遗传信息的传递机制
转录和翻译的基本概念
基因信息的表达遵循被称为“中心法则”的核心理论:DNA → RNA → 蛋白质。也就是说,遗传信息从DNA流向RNA,再流向蛋白质,由此实现遗传性状的表现。这个过程中包括两个至关重要的步骤:转录和翻译。
转录(Transcription)
在转录过程中,细胞利用DNA的其中一条链作为模板,合成出一条互补的RNA分子。这个过程主要由RNA聚合酶催化完成。在大多数基因中,转录产物主要是信使RNA(mRNA)。mRNA分子类似“信使”,把DNA上携带的遗传指令带出细胞核,成为后续翻译过程的模板。除了mRNA,转录还会产生一些其他类型的RNA(如rRNA、tRNA等),但它们的功能不同。在真核细胞中,转录后的初级mRNA(前体mRNA)还需要经过剪接、加帽和加poly-A尾等多种加工步骤才能成为成熟的mRNA。
翻译(Translation)
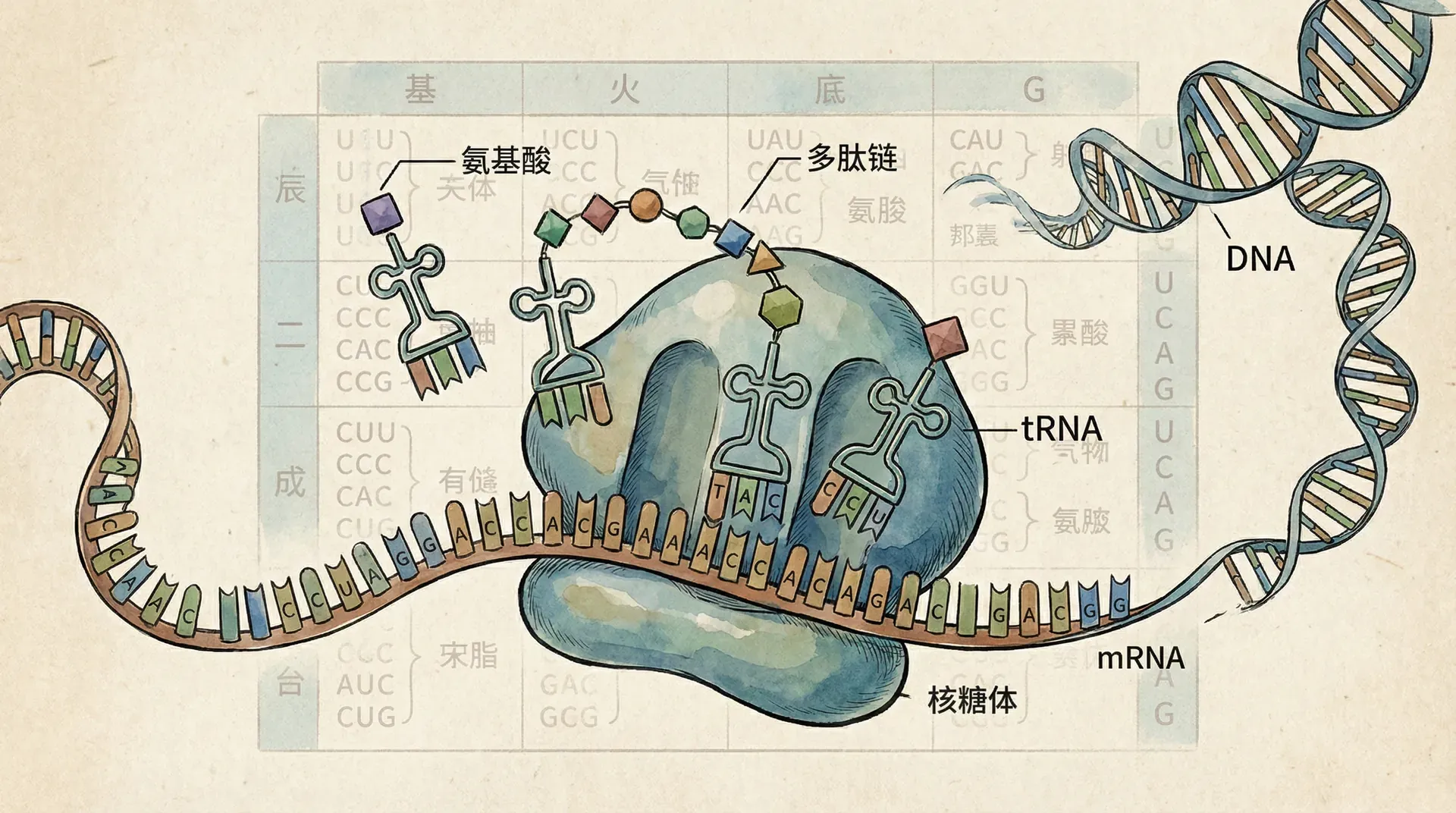
翻译是遗传信息由RNA分子转变为蛋白质的过程。在细胞质中,成熟mRNA与核糖体结合,核糖体作为蛋白合成的“工厂”,负责读取mRNA上的遗传密码(每组三个核苷酸为一组,称为“密码子”),并据此合成特定序列的氨基酸链。在这一过程中,转运RNA(tRNA)发挥“翻译器”的作用。每种tRNA分子能够识别特定的mRNA密码子,并携带相应的氨基酸。“反密码子”片段确保tRNA与正确的mRNA密码子配对,实现氨基酸的正确顺序连接,最终形成具有生物学功能的蛋白质。
在此过程中,“中心法则”不仅揭示了遗传信息如何沿着单向途径传递,还暗示了生物学系统对遗传信息流的高效和精确调控。实际上,基因的表达还受到复杂的调控,包括启动子的激活/抑制、mRNA的选择性剪接、翻译效率调节等,这种多层次调控保证了细胞能够根据内外部环境的变化灵活地调节蛋白质的合成。
原核生物与真核生物的区别与联系
在原核生物(如大肠杆菌)中,细胞结构相对简单,遗传物质并没有被核膜包裹。因此,转录和翻译通常可以在同一空间、几乎同时进行——有时mRNA还未合成完毕,翻译过程就已在其5'端开始。这一同步性有利于原核生物快速响应环境信号与资源变化。而且,原核生物的mRNA一般没有复杂的加工修饰,寿命较短。
相比之下,真核生物(如人类和高等植物)具有高度分化的细胞结构。它们的DNA被封闭在细胞核内,转录过程仅能在核内进行。生成的前体mRNA必须经过“加帽”“切除内含子加连接外显子(剪接)”“加poly-A尾”等一系列加工和修饰,最终形成成熟mRNA,才能穿过核孔进入细胞质参与翻译。这样的分隔和严格加工过程提高了表达调控的精度,但也使基因表达的整个流程更加复杂和多样。
此外,真核生物还拥有复杂的基因组结构,例如存在大量的内含子、调控元件以及染色质结构对基因表达产物的影响。不仅成千上万的基因需要被有序和有选择地表达,而且这种表达在不同的组织、发育阶段和环境条件下都可能不同,展现出高度的空间和时间特异性。
总之,虽然原核与真核生物的遗传信息传递基本遵循同一“中心法则”,但在结构和调控机制上展现了显著差异。这些差异体现了生物进化过程中复杂性的不断增强,也反映了生命活动高度的精确性与灵活性。
遗传密码的奥秘
密码子的基本特征
遗传密码是指存在于DNA或RNA分子中的一套信息编码体系,它由每组三个连续碱基(即一个密码子)来指代一种氨基酸或一个翻译信号。我们的DNA和RNA中存在4种不同的碱基——腺嘌呤(A)、胞嘧啶(C)、鸟嘌呤(G)、胸腺嘧啶(T)/尿嘧啶(U),每三个为一组,因此理论上三联体密码能组合成4³ = 64种不同的密码子。这些密码子承担着翻译遗传信息、指导细胞合成蛋白质的重要使命。虽然仅有20种标准氨基酸参与蛋白质的构建,但冗余的密码子数量确保了遗传信息传递的稳定性与容错性。
遗传密码高度保守且几乎通用于地球上的所有生物,从最原始的细菌到高级的动植物甚至人类,无例外地遵循着这套编码。这一现象也被称为“遗传密码的普遍性”,是支持不同生物具有共同起源、说明生命演化连续性的有力科学证据。
遗传密码的一大特点还在于其“无重叠”和“无逗号”——即密码子之间不重叠,每组三个一读,不跳过、不重复,这为蛋白质序列的准确表达打下了基础。此外,遗传密码的起始位置决定了后续全部的三联体划分,因此,起始密码子的识别至关重要,否则会导致移码突变(frame-shift),进而造成后续蛋白质的功能异常。
密码子的功能分类
值得注意的是,遗传密码具有显著的简并性(即冗余性):部分氨基酸由多个不同的密码子编码。例如,亮氨酸有6个密码子,精氨酸和丝氨酸也各有6个密码子。这种冗余可以有效降低点突变对蛋白质功能的潜在危害,提高了生命体系的信息稳定性。另一方面,每个密码子只对应一种氨基酸,则确保了翻译过程的单一性和准确性,避免了歧义现象的发生。这种独特的编码规律为生物遗传信息的高效、准确传递提供了分子基础。
密码子的这一特性,还使科学家在蛋白工程、基因编辑等领域能通过设计性变异,影响或优化蛋白表达,提高合成生物学的操作能力。
虽然大多数情况下遗传密码表是统一的,但自然界中某些生物和细胞器(如线粒体)偶尔会有少数密码子的“重新分配”。比如人体线粒体中的UGA可编码色氨酸而不是标志终止,UAA、AUA等也可能有不一样的指令。这些例外为分子进化研究提供了深刻的信息。
中国科学家对遗传密码的贡献
在遗传密码破译的国际竞争中,中国科学家同样做出了不可忽视的贡献。早在1965年,中科院上海生化所的科学家团队成功全合成并结晶获得了具有生物活性的牛胰岛素分子,这是人类首次实现人工合成完整蛋白质,为人类解读蛋白质的结构与遗传信息的关系奠定了基础。此外,国内学者参与了密码子、mRNA序列与蛋白质序列间“翻译程序”的研究,也为我国后续分子生物学、遗传工程、基因药物等学科的发展打下了坚实基础。
RNA的加工与成熟
真核生物mRNA的修饰
在真核细胞中,转录产生的并非可以直接参与翻译的成熟mRNA,而是一段称为前体mRNA(pre-mRNA)的原始转录产物。为了成为功能完善的mRNA并导出细胞核,pre-mRNA在加工和成熟过程中经历三大关键步骤,使其具备稳定性、高效性,并可被核糖体正确识别。
1. 5'端加帽(5' Capping)
在mRNA的5'端,通过一系列酶促反应加上一个7-甲基鸟嘌呤“帽”结构。这不仅能保护mRNA免遭核酸外切酶降解,还参与核输出、翻译起始复合物的组装等关键环节。加帽结构几乎是所有真核mRNA的专有标记。
2. 3'端多聚腺苷酸化(Polyadenylation)
mRNA的3'端并不是转录终止后自然结束的,而是借助聚腺苷酸化酶在其末尾添加50-250个腺嘌呤核苷酸构成的poly-A尾。“尾巴”一方面增强了mRNA的稳定性和半衰期,另一方面参与mRNA核输出并促进翻译。Poly-A尾的长度及调控还对基因表达强度和蛋白质亚型产生深远影响。
3. RNA剪接(Splicing)
最复杂的环节莫过于RNA剪接。真核基因通常由编码区(外显子)和非编码区(内含子)交替排列组成。在剪接过程中,含有内含子的pre-mRNA在剪接体(spliceosome)等分子机器作用下精准切除内含子,并将多个外显子首尾连接,生成结构连续、可翻译的成熟mRNA。外显子最终决定了蛋白质的氨基酸序列。
此外,真核细胞mRNA的加工还包括RNA编辑(个别碱基发生改变)、RNA定位等调控机制,这让基因表达的精确性和多样性进一步提升。
选择性剪接的生物学意义
所谓“选择性剪接”(Alternative Splicing),是指在同一个基因的前体mRNA上,可以通过不同的方式组装外显子,排除不同内含子片段,从而生成多种不同的mRNA分子,最终翻译成功能或结构各异的蛋白质。选择性剪接是复杂生物重要的遗传多样性来源。
人类的一个基因经过选择性剪接后,可以在神经细胞中生成一种蛋白,在肌肉细胞中生成另一种蛋白,这赋予了多细胞生物极高的适应性。选择性剪接机制也解释了为何人类基因组仅有约2万个蛋白编码基因,却能够合成超过10万种不同蛋白质。
若RNA剪接环节发生错误或调控失常,往往会导致严重的遗传疾病。例如,β-地中海贫血的一些类型,就是因β-珠蛋白基因的RNA剪接异常,产生功能缺失的血红蛋白,从而出现贫血症状。其他如脊髓性肌萎缩症等疾病也与选择性剪接异常有关。
翻译过程的分子机制
tRNA的结构和功能
tRNA(转运RNA)是连接遗传信息与蛋白质合成的关键适配分子。它的结构呈“苜蓿叶”状,在空间中折叠为“L”形。每个tRNA有两个主要功能区域:
- 氨基酸结合臂:3'末端能特异性结合一种氨基酸。每种氨基酸都至少有一种相应的tRNA配对。
- 反密码子环:tRNA底部的一个环结构,包含一个三联体反密码子(anticodon),能与mRNA上的密码子以碱基互补配对。这样确保正确的氨基酸被放置到正在合成的多肽链相应位置。
在蛋白质合成过程中,氨基酸首先被“氨基酰-tRNA合成酶”准确地加载到对应的tRNA上,不同的氨基酰tRNA合成酶专一识别各自的tRNA和氨基酸。这一反应的高保真性对于蛋白质合成的正确进行至关重要。
蛋白质合成的基本步骤
蛋白质翻译过程在细胞质的核糖体上完成,通常包括以下三个经典阶段:
除了常规的翻译步骤外,真核与原核生物在翻译调控与效率上各有特色。例如真核细胞的mRNA常常在形成“多核糖体”(polysome)结构的状态下被同时多个核糖体翻译,大大提高了蛋白合成的效率。
基因表达的生物学意义
蛋白质多样性的来源
生命复杂性的一大根源在于有限的基因数量却能决定极其丰富的蛋白质种类。基因表达系统通过多种方式实现蛋白质的多样化,例如:
选择性剪接显著拓展了蛋白供给的方式,使一种DNA序列指导的前体mRNA通过不同的选择、剪切和拼接产生多种成品mRNA,从而翻译为功能差异巨大的一组蛋白。翻译后修饰则指多肽链合成完成后,还需经由磷酸化、甲基化、糖基化等修饰,使蛋白质拥有独特的活性、定位和相互作用能力。而蛋白质折叠带来结构各异的空间构型,为蛋白质多样性与功能性添加更高维度的变化。
基因表达调控的重要性
基因表达的调控贯穿生命全周期,从单细胞分化、器官形成到各种环境响应,无不依赖于基因表达的时空精准调节。例如在胚胎发育过程中,细胞需精确在正确的时间、部位激活适当的基因,才能保障组织器官的正常分化和功能形成。基因表达也因环境刺激、信号因子的调节而灵活应变。
遗传性疾病、癌症、自身免疫等众多疾病都与基因表达异常有关,因此基因治疗、mRNA疫苗、个性化医疗等新兴技术正是基于分子层面对基因表达的精准操控和深入认识。只有全面剖析从基因到蛋白质的信息流动、调控及异常机制,我们才能揭示生命现象的核心秘密,研发更有效的诊断与治疗手段,为人类健康和社会发展提供坚实的科学支撑。
核糖体
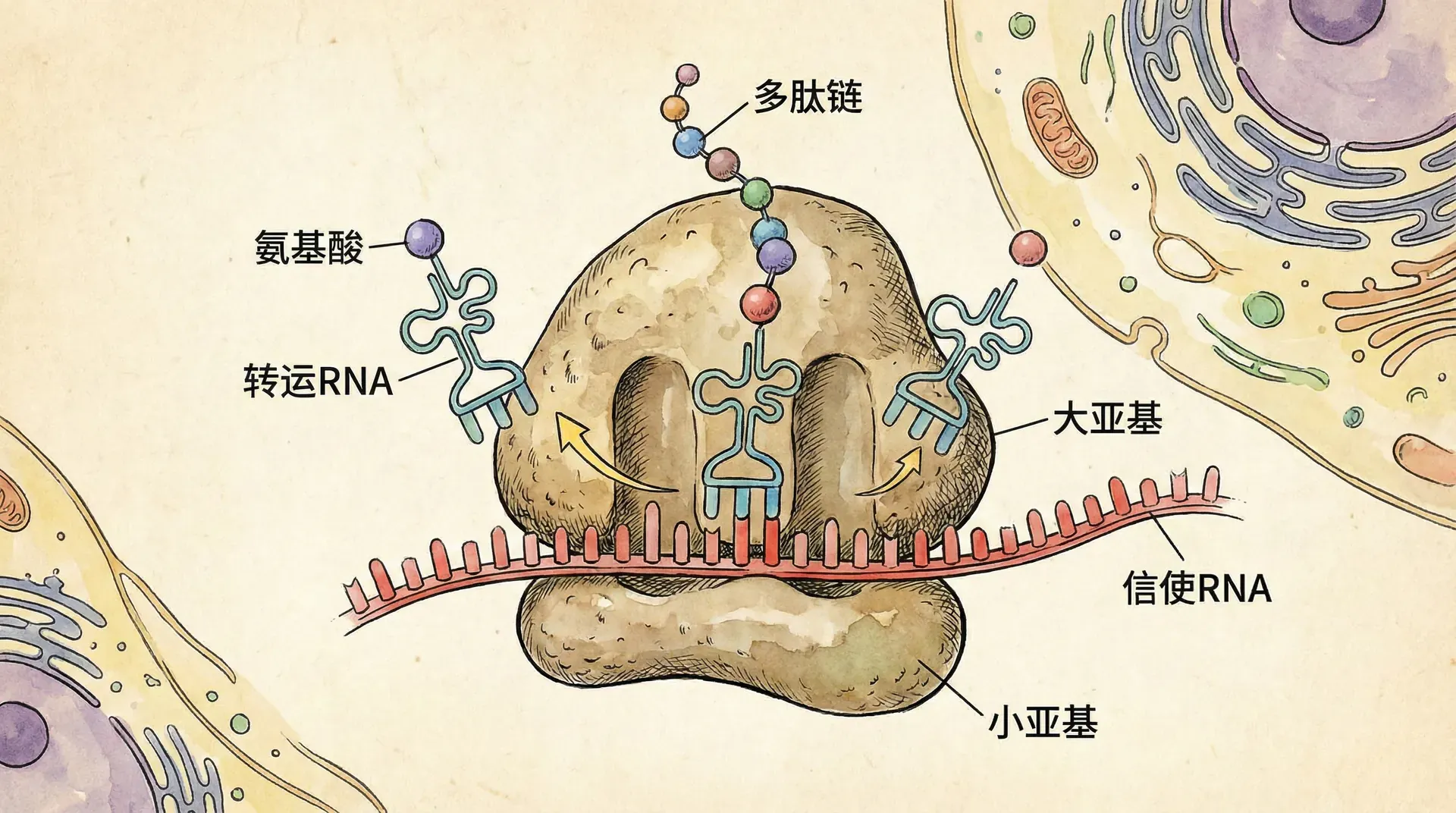
核糖体的基本结构
核糖体是所有活细胞中负责蛋白质合成的复杂分子机器,由RNA和蛋白质组成。核糖体具有精巧的结构组织,包括大小两个亚基。在原核生物中,核糖体为70S型(包含30S小亚基和50S大亚基),而真核生物的核糖体为80S型(包含40S小亚基和60S大亚基)。
核糖体的核心功能在于其三个重要的结合位点:
核糖体RNA的催化作用
现代分子生物学研究表明,核糖体中的rRNA(而非蛋白质成分)是肽键形成的真正催化者。这一发现颠覆了人们对酶催化的传统认识,证明了RNA分子同样具有催化活性,核糖体本质上可以被视为一个巨大的核酶(ribozyme)。
rRNA分子在核糖体内部形成复杂的三维结构,其中大亚基的rRNA负责催化肽键的生成。这种RNA催化活性可能反映了早期“RNA世界”的遗迹,暗示在蛋白质出现之前,RNA分子就已经承担着重要的生物催化功能。
核糖体蛋白质主要分布在核糖体的外表面,它们的主要作用是维持rRNA的正确构型,协助rRNA发挥催化功能,而不是直接参与肽键的形成。这一发现进一步证实了rRNA在蛋白质合成中的核心地位。
蛋白质的转运与定位
信号肽介导的蛋白质靶向
细胞内蛋白质的正确定位对其功能发挥至关重要。许多蛋白质携带特定的信号序列,指导它们运输到适当的细胞区室。其中最重要的是内质网信号肽,通常位于新合成多肽链的N端前20个氨基酸左右。
当携带内质网信号肽的多肽链从核糖体上伸出时,立即被信号识别颗粒(SRP)识别并结合。SRP是一个由蛋白质和RNA组成的核蛋白复合体,能够暂停翻译过程,并引导整个翻译复合体移动到内质网膜表面。
蛋白质的协同转运
在内质网膜上,SRP与SRP受体结合,形成蛋白质转运复合体。核糖体重新开始翻译,新生的多肽链一边合成一边通过转运子穿过内质网膜,进入内质网腔。这种“协同转运”机制确保了跨膜蛋白和分泌蛋白的正确加工和折叠。
对于其他细胞器(如线粒体、叶绿体、细胞核)的蛋白质,则采用不同的信号肽和转运机制。通常这些蛋白质在细胞质中完成翻译后,再通过特异性的转运机器进入目标细胞器。
翻译后修饰的生物学功能
新合成的多肽链往往需要经历多种翻译后修饰才能获得完整的生物学功能。这些修饰包括:
这些修饰不仅丰富了蛋白质的功能多样性,还为细胞提供了动态调节蛋白质活性的重要机制。
基因突变对蛋白质功能的影响
点突变的分子后果
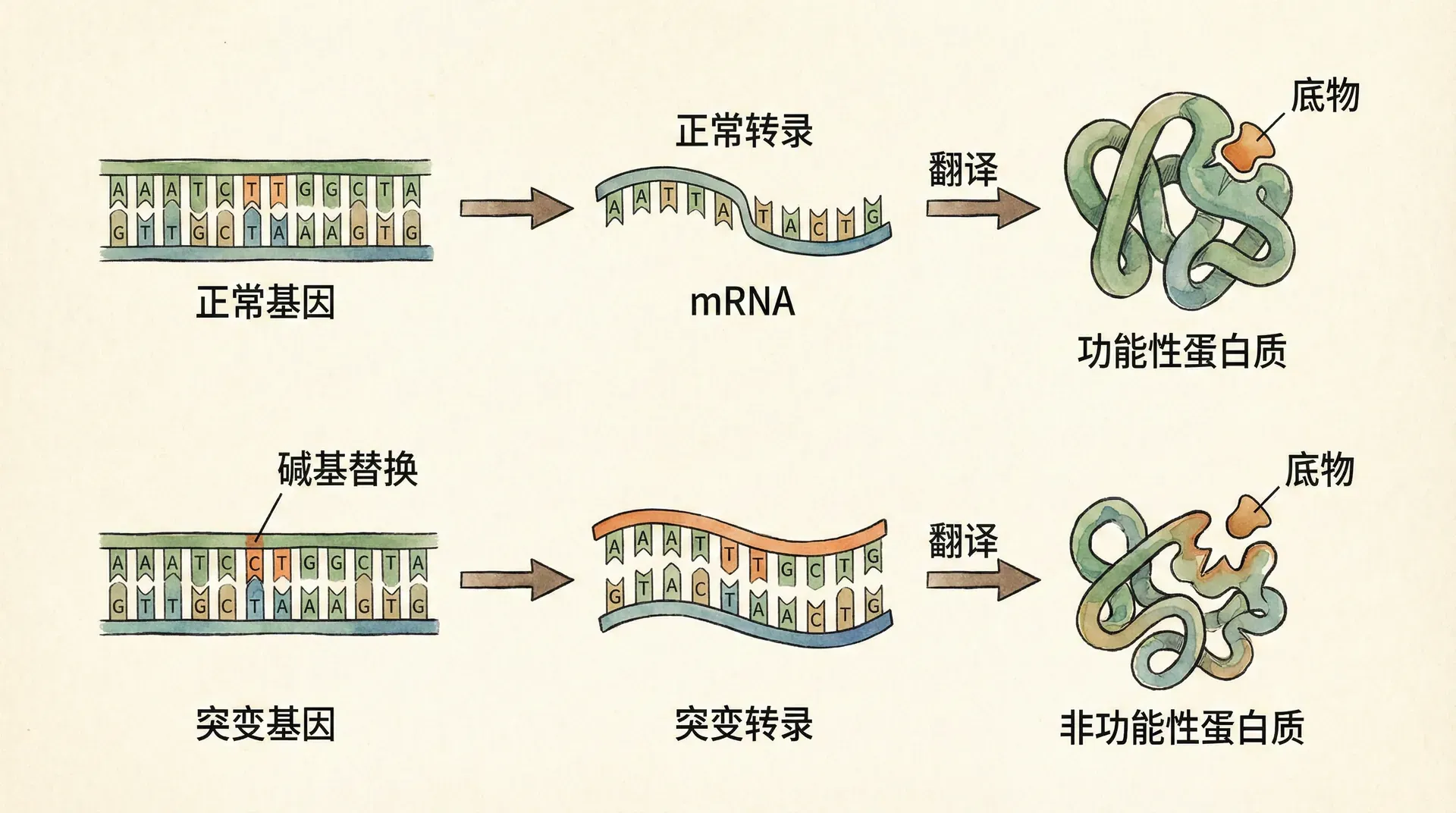
基因序列中单个核苷酸的改变可能导致截然不同的生物学后果。根据对蛋白质序列影响的不同,点突变可以分为几个主要类型:
静默突变:由于遗传密码的简并性,某些核苷酸变化并不改变对应的氨基酸,这类突变通常对蛋白质功能无明显影响。然而,最新研究表明,即使是静默突变也可能通过影响mRNA的稳定性、翻译效率或蛋白质折叠动力学而产生微妙的表型效应。
错义突变:导致氨基酸改变的突变,其后果取决于新氨基酸的理化性质以及该位置在蛋白质结构中的重要程度。发生在蛋白质活性位点或关键结构域的错义突变往往严重影响蛋白质功能。
无义突变:将编码氨基酸的密码子改变为终止密码子,导致翻译提前终止,产生截短的、通常无功能的蛋白质。
镰刀型细胞贫血症的分子机理
镰刀型细胞贫血症是点突变致病的经典例子。β-珠蛋白基因的第6个密码子从GAG(编码谷氨酸)突变为GTG(编码缬氨酸),这一氨基酸的替换看似微小,却产生了严重的病理后果。
谷氨酸是亲水的酸性氨基酸,而缬氨酸是疏水的非极性氨基酸。这种性质的改变使得血红蛋白分子在缺氧条件下倾向于相互聚集,形成刚性纤维,导致红细胞变形为镰刀状,引发血管阻塞和组织缺氧等一系列临床症状。
镰刀型细胞贫血症的例子说明了蛋白质结构与功能的密切关系。即使是单个氨基酸的改变,如果发生在关键位置,也可能完全改变蛋白质的生物学特性,凸显了基因序列精确性的重要意义。
框移突变的严重后果
插入或缺失突变(特别是非3的倍数的核苷酸变化)会导致框移突变,这是最具破坏性的突变类型之一。框移改变了突变点下游所有密码子的读码框,通常很快遇到终止密码子,产生截短且无功能的蛋白质。
框移突变的严重性在于其影响范围广泛。即使是单个核苷酸的插入或缺失,也会影响该位点之后的全部氨基酸序列,除非框移发生在基因的最末端,否则几乎必然导致蛋白质功能的完全丧失。
突变与疾病的关系
现代医学研究表明,许多遗传性疾病都可以追溯到特定的基因突变。通过深入理解基因突变如何影响蛋白质结构和功能,科学家们正在开发精准的基因治疗策略,为这些疾病的治疗带来新的希望。
总结
遗传密码在地球上所有生命形式中的高度保守性,为我们理解生命的共同起源提供了强有力的证据。这种普遍性不仅体现了早期生命演化过程中信息编码系统的固化,也反映了任何对遗传密码的重大改变都可能带来灾难性后果的选择压力。
然而,在某些极端环境中的生物(如深海热泉细菌)以及细胞器(如线粒体和叶绿体)中,确实存在遗传密码的变异。这些例外为我们研究密码演化和适应性变化提供了珍贵的自然实验材料。
对基因表达机制的深入理解,为现代生物技术的发展奠定了坚实基础。mRNA疫苗技术的成功应用,基因编辑技术的精确操作,以及合成生物学的蓬勃发展,都直接受益于我们对转录翻译过程的精确认识。
未来,随着对基因表达调控网络理解的不断深化,我们有望在疾病治疗、农作物改良、生物能源开发等多个领域实现更大突破。基因表达的研究不仅帮助我们理解生命的本质,更为解决人类面临的重大挑战提供了强有力的工具。