生态学野外实验

在自然界中,生物之间的相互作用错综复杂。比如一片植物,影响它们生长和繁衍的因素可能有很多:土壤中的肥力和养分、降雨的多少、气温的冷暖,还要面对食草动物的啃食、病虫害的侵袭,以及周围其他植物与它们争夺阳光和空间。这些因素往往不是单独起作用,而是彼此交织、相互影响,造成结果的变化莫测。对于生态学家来说,理解这些因素各自的作用以及它们如何协同、冲突,是解开自然规律的重要一步。
那么,如何在这样纷繁复杂的环境中分清每个因素的影响?面对无法在实验室里彻底控制变量的现实,生态学家们的答案,是设计并实施精心策划的野外实验。通过野外实验,可以在人为可控的前提下,观察和分析环境、物种及它们之间关系的细微变化,从而一步步揭示大自然背后的内在规律。
如何设计一个科学的生态实验

设计一个可靠的生态实验并不简单。假设你想知道某种鸟类对森林昆虫数量的影响。最直接的想法是找一片森林,把鸟赶走,然后观察昆虫数量的变化。但这样做够科学吗?
这里有个关键问题:即使你发现昆虫数量增加了,你怎么知道这是因为鸟类减少,而不是因为那段时间恰好天气变暖、降雨增多,或者其他什么原因呢?自然环境每时每刻都在变化,温度在波动,湿度在改变,各种生物也在迁徙流动。
对照组是生态实验的核心。我们需要在相同的环境中设置两片区域:一片是实验区(比如驱赶鸟类),另一片是对照区(保持自然状态)。只有这样,才能确保观察到的差异真正来自我们的操作,而不是环境的自然波动。
但仅有对照组还不够。如果你只设置了一个实验区和一个对照区,而实验区恰好在山坡的阳面,对照区在阴面,那么两者的差异可能只是因为光照不同,而非鸟类的作用。因此,我们需要在空间上随机分布多个实验区和对照区,这样才能排除位置因素的干扰。
另一个容易犯的错误叫做“伪重复”。比如,你在一片实验区里设置了10个观测点,然后把这10个点当成10次独立实验。但实际上,这10个点之间距离很近,共享着相同的微环境、相同的种群,它们并不是真正独立的。这就像你测量同一个人的身高10次,不能说你测量了10个人。真正的重复是指在不同的地点、不同的时间,独立进行多次实验。
生态实验还有一个难点:结果往往出乎意料。你原以为移除某种捕食者会让被捕食者大量繁殖,结果却发现整个群落发生了连锁反应,最终的结果完全不是你预期的那样。这种“间接效应”在生态系统中非常普遍,也让生态实验的解读变得格外困难。
捕食者如何维持生物多样性
现在让我们通过一个真实的研究来理解生态实验的价值。这个实验关注的是鄱阳湖湿地系统中,候鸟对底栖生物多样性的影响。
鄱阳湖是中国最大的淡水湖,每年冬季有数十万只候鸟在此栖息,其中包括大量的涉禽,它们以湖底的螺类、贝类等软体动物为食。研究人员想知道:这些鸟类的捕食活动,对湖底生物群落有什么影响?
实验设计是这样的:在湖滨湿地选择几块相似的区域,用网罩隔离其中一些区域,防止鸟类进入,而让其他区域保持自然状态。两年后,研究人员统计了各个区域的底栖动物物种数量。结果令人惊讶:
有鸟类的区域反而维持了更高的物种多样性。这是为什么呢?
原来,在湖底的空间争夺战中,有一种大型田螺特别强势,它们繁殖快、个体大,能够占据大量的优质栖息地。但这种田螺恰好是鸟类最喜欢的食物。当鸟类不断捕食这些优势种时,它们为其他较弱的物种腾出了生存空间。那些小型螺类、幼贝等虽然竞争力弱,但能迅速在鸟类制造的空白地带定居下来。
这种通过控制优势种而维持群落多样性的物种,被称为“关键种”。它们在生态系统中扮演着超乎其数量的重要角色,一旦失去,整个群落结构可能发生剧变。这个发现对湿地保护有重要启示。保护候鸟不仅是为了鸟类本身,也是在维护整个湖泊生态系统的健康和多样性。
物理环境与生物竞争的博弈
生物的分布不仅受到其他生物的影响,物理环境的限制同样重要。太湖的一项研究很好地展示了这两种力量如何共同塑造生物的分布格局。
太湖是一个浅水湖泊,湖岸边有大量石块和混凝土结构。在这些硬质表面上,生长着两种常见的附着藻类:绿藻和硅藻。有趣的是,这两种藻类在垂直方向上有着清晰的分界:绿藻主要分布在水面附近,硅藻则占据着较深的水下区域。
研究人员做了一系列实验来探究这种分布模式的原因。他们在不同水深处放置新的基质(干净的石块),观察藻类的定居和生长情况。结果发现,两种藻类的幼孢子其实都能在各个深度附着和萌发,说明它们的分布范围原本可以更广。
那是什么限制了它们的分布呢?进一步的实验给出了答案:
在浅水区,当研究人员人为清除绿藻后,硅藻能够迅速扩张到浅水区域,生长良好。这说明在浅水区,硅藻是输给了绿藻的竞争,而不是环境不适合它生长。绿藻生长迅速,能够快速覆盖基质表面,遮挡光线,硅藻便无法立足。
但在深水区,情况完全不同。即使清除了硅藻,绿藻也无法在深水区长期存活。原因很简单:深水区光照弱、水流冲刷力强,超出了绿藻的耐受范围。而硅藻的细胞壁含有硅质,更加坚固,能够适应深水的严苛条件。
这个研究揭示了一个普遍规律:在垂直分布的梯度上,生物分布的上限往往受物理环境限制,而下限则主要受生物竞争决定。绿藻虽然竞争力强,但承受不了深水的物理压力;硅藻虽然耐受力好,但在浅水区竞争不过绿藻。
竞争也会因时而变
生态学中的竞争并不是一成不变的,环境条件的改变会显著影响竞争的强度。秦岭南北坡的一项长期研究展示了这一点。
秦岭是中国重要的地理分界线,南坡北坡的气候差异明显。研究人员在秦岭山地选择了两种常见的小型食虫鸟类:大山雀和褐头山雀。这两种鸟的食性相似,都以树上的昆虫为主要食物,存在潜在的食物竞争。
实验持续了四年,其中两年降雨充沛(湿润年),另外两年降雨偏少(干旱年)。研究人员在一些样区中移除了体型较大的大山雀,观察小型的褐头山雀的响应。
数据清楚地表明,在湿润年份,森林中昆虫资源丰富,即使两种鸟共存,食物也足够分配,竞争并不明显。但到了干旱年份,昆虫数量锐减,食物成了稀缺资源,竞争才真正激烈起来。
更有意思的是,竞争的影响是不对称的。移除大山雀对褐头山雀产生了多方面的积极效应,但反过来移除褐头山雀,对大山雀的影响却微乎其微。这说明在这个系统中,大山雀处于竞争优势地位,它对小型褐头山雀有压制作用,而褐头山雀对它几乎构不成威胁。
这项研究提醒我们,在评估物种间的相互作用时,不能只看一个时间点。环境的年际波动会让生物间的关系呈现出完全不同的面貌。这对于预测气候变化对生态系统的影响尤其重要。
草原上的复杂关系网
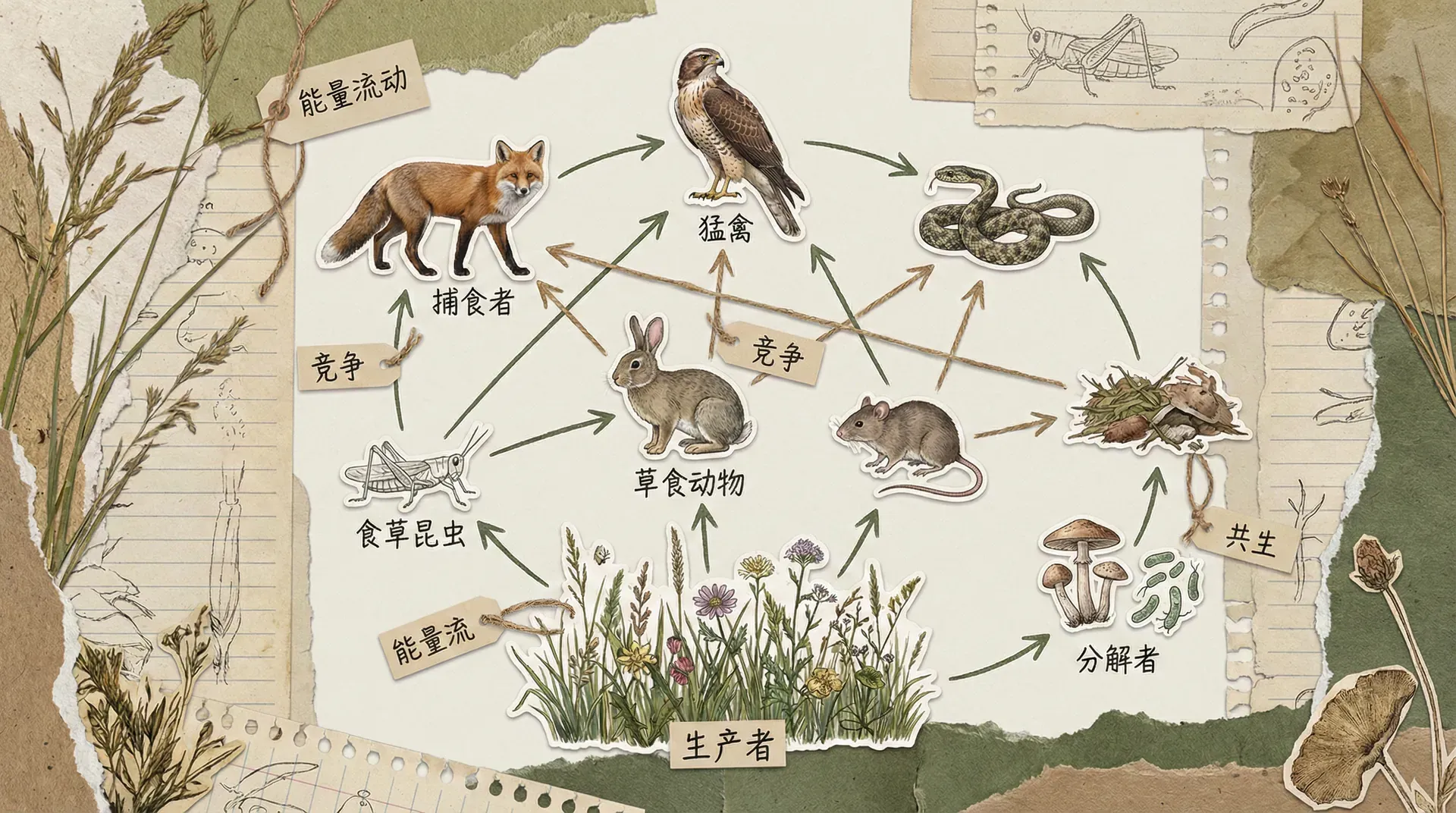
生态系统中的相互作用远比我们想象的复杂。内蒙古锡林郭勒草原的一项大规模实验,展示了种子、啮齿动物和蚂蚁之间错综复杂的关系。
这个实验的设计相当精密。研究团队在草原上划出了24个50米×50米的样方,用围栏把每个样方隔开。围栏底部有大小不同的孔洞:有的孔洞小(直径2厘米),只允许小型鼠类如小仓鼠通过,而挡住大型的跳鼠;有的孔洞大(直径7厘米),所有啮齿动物都能自由进出。通过活体捕捉和药物处理,研究人员能够控制特定样方中的动物组成。
实验处理包括多种组合:
这个实验持续了长达8年,积累了大量数据。短期结果看起来很直观:啮齿动物和蚂蚁确实在竞争种子。移除老鼠的样方中,某种小型蚂蚁的数量明显增加;反过来,移除蚂蚁的样方中,老鼠的数量也有所上升。
但随着实验的继续,一个意想不到的模式浮现出来。研究人员发现,从长期来看,啮齿动物的存在实际上对蚂蚁有利。怎么会这样呢?
答案藏在植物群落的变化中。草原上的植物有大粒种子和小粒种子之分。啮齿动物偏好大粒种子,蚂蚁则更喜欢小粒种子。当啮齿动物大量取食大粒种子时,会抑制大籽植物的繁殖,给小籽植物腾出生长空间。而小籽植物的繁盛,反过来为蚂蚁提供了更丰富的食物来源。
这是一个典型的“间接促进”案例。两个物种在直接层面上是竞争关系(都吃种子),但通过影响第三方(植物群落结构),一方的存在最终让另一方受益。生态学家把这种通过中间环节产生的效应称为“间接相互作用”。
这项研究告诉我们,仅仅观察两个物种之间的直接相互作用是不够的,必须把整个群落的关系网络都考虑进来。这也解释了为什么许多生态实验会得到出乎意料的结果。
自然恢复的奥秘
生态系统在遭受破坏后能否自我恢复?恢复需要多长时间?这些问题对于生态保护和修复工作至关重要。西双版纳热带雨林的一项实验提供了启发性的答案。
研究团队在西双版纳保护区选择了几块小型林地(每块约100平方米),这些林地彼此隔离,就像小岛一样。研究人员用药物熏蒸的方法,彻底清除了这些林地中的所有节肢动物(昆虫、蜘蛛等),但植被基本保持完整。然后,他们开始监测这些林地的重新定居过程。
节肢动物的回归速度超出了预期。实验开始后的最初几周,物种数量快速攀升,各种昆虫、蜘蛛从周围的森林迁入这些空白区域。大约200天后,物种数量趋于稳定,达到了一个相对平衡的状态。
但仔细分析会发现,恢复后的群落与原来的群落并不完全相同。某些林地的物种数量略低于原始水平,群落的物种组成也有所不同。这可能有两个原因:一是新迁入的物种之间的竞争关系比原来更激烈,相互干扰更多;二是新移民对当地资源的利用效率不如原住民。
研究人员还统计了不同营养级的物种数量变化:
从这个表格可以看出,各个营养级的物种数量虽有波动,但大体上回到了相近的水平。食草类昆虫、蚂蚁、捕食类的物种数基本恢复,维持了群落的基本结构。这暗示着生态系统可能存在某种内在的组织规律,使得不同营养级之间保持相对稳定的比例关系。
这个实验带来了乐观的信息:在周围有健康生态系统作为种源的情况下,受损的小型栖息地能够在相对较短的时间内实现物种的重新定居。但我们也要意识到,群落的功能恢复可能比物种数量的恢复要慢得多,完全恢复到原始状态可能需要更长的时间。
总结
通过这些野外实验,我们逐渐理解了生态系统运作的一些基本规律。关键种的存在维持着群落的多样性,物理环境和生物竞争共同塑造着物种的分布,竞争强度随着环境条件波动,物种间存在复杂的间接相互作用,受损的生态系统具有一定的自我修复能力。
这些发现不仅增进了我们对自然的认识,也为生态保护和恢复提供了科学依据。保护关键物种、维护栖息地的异质性、关注气候变化带来的资源波动、理解物种网络而非孤立的种对关系,这些都是实验生态学留给保护工作的启示。
当然,野外实验也有其局限性。生态系统太过复杂,我们的实验规模总是有限的,实验周期相对于生态过程也往往太短。许多重要的生态问题,如物种的长期演化适应,很难通过短期实验来回答。但正是这些精心设计的实验,让我们能够在纷繁复杂的自然现象中,识别出因果关系,理解生态学的基本原理。
大自然是一本厚重的书,而野外实验是我们读懂这本书的重要工具。每一个实验都像一个探针,帮助我们窥见生态系统的运作机制。随着更多实验的开展,我们对自然的理解将会越来越深入,保护和恢复生态系统的能力也将越来越强。