德尔布吕克模型
生命进化的漫长历史中,科学家一直在不断追问一个根本性的问题:为何遗传信息能够在亿万年间保持惊人的稳定性,同时又具备适应环境、引发变异和创新的能力?这背后的奥秘,德尔布吕克的分子遗传学理论给出了物理学层面的深刻洞见。他用物理和化学的视角,将遗传的守恒与变异归结为一种微妙而动态的平衡机制,使我们能够真正理解,何以生命既能“守旧”、维持遗传信息的高度稳定,又能“创新”,在特定条件下产生遗传变异,推动物种的演化和多样性。
分子层面的遗传密码
在物理学家的眼中,基因并不是一种死板凝固的结构。它是动态的、充满活力的高分子体系。基因分子(如DNA)的本质,其实更接近一种微观级别上的复杂机器:它的基本组成单位和整体化学成分可以长期保持不变,但在内部,每一个原子的位置都有可能在极小的尺度中发生跳跃和重排。这些跳跃和重排可能导致基因分子形成不同的构型(同分异构体),即便外表上看分子式没有变化,其三维空间结构和功能状态却可能大不一样。
德尔布吕克模型阐明,基因结构的改变通常不是像水流一般缓慢、连续地发生,而是类似于“隧穿”或者“跨能垒跳跃”的突发事件。这些微小的结构变化,正是生命多样性的物质基础。令人惊奇的是,虽然基因内部的排列存在着多种潜在的可能性,但实际上绝大多数原子排列都牢牢处于能量最低态,很少发生“自发地”改变。
值得强调的是,即便是极微小的分子层次构型变化,也能引起巨大的生物学差异,这正是生命系统极其敏感与多样的根本原因。但要实现这些微小变异,大多数情况下需要突破高能量障碍(能垒),这大大降低了突变的发生概率。因此,在分子世界中,基因突变实际上是一种极其稀有的事件,这为遗传信息的长期稳定性提供了保障。
我们可以将基因分子的稳定性类比为坚不可摧的万里长城:想要让整体结构保持稳固,每一块城砖(原子或分子单元)都必须正确安置、严丝合缝。在分子尺度上,只有当系统获得了足够的能量,原子才能从一个稳定位置“翻越”高能垒跳到另一种排列。在一般的生理条件下,这样的高能障碍让原子难以任意移动,因此保证了基因在细胞分裂和后代传递过程中发生差错的概率极低。
然而,正如德尔布吕克模型所强调的那样,虽然单一分子的跃迁极为罕见,但考虑到生命体中存在着亿万个细胞、长时间的世代更替,即便极小概率的突变也会不断积累,从而推动物种的进化。这一理论的重要意义在于,它揭示了遗传现象受制于能量壁垒(物理稳定性)和统计概率(大数法则)双重机制,科学地解释了遗传稳定与进化动力之间的统一和张力。
量子理论框架下的分子键合
那么,为什么要诉诸量子力学来理解生命的分子基础?这个问题的重要性不亚于我们运用材料科学去解析青藏铁路能够穿越极端高原地形的奥秘。量子力学是人类历史上第一次用基本的数学与物理原理,从最微观的粒子出发,成功预测和解释了原子的聚集方式、化学键合乃至分子的复杂行为,并由此揭示了自然界的底层规律。
尤其值得强调的是,海特勒-伦敦共价键理论的提出具有划时代意义。这一理论并不是生硬为了化学服务,而是追寻量子力学深层次原理的副产品。令人震撼的是,这个纯理论演算推导出的结果,恰恰能够精确吻合实验上观察到的化学键结构与能量变化。正如德尔布吕克所言,“这样理论与实验的完美契合,此后在量子理论发展史中罕有出现”,突显了这一理论在科学史上的特殊地位。
量子理论不仅解释分子的结构稳定性,还让我们明白:为什么有的分子几乎不可能自发发生结构上的巨大跳跃。能量量子化限制了分子内部原子的运动轨迹和跃迁模式,使得大多数分子能长时间保持其结构。在生命体内,这一机制为基因的稳定和高效信息传递提供了坚实的物理基础。
物质聚集态的新分类体系
传统物理和化学的分类体系通常将“分子”、“固体”、“晶体”等视为完全不同的物质形态。这种粗略的划分,如同我们仅以行政区划来理解中国疆域上的山川湖泊那样,虽然有助于快速记忆,但很容易掩盖真正的物理本质。现代量子理论则给出了全新的聚集态本体论分类:
这种重新分类的意义如同重新审视中华文明:看似完全不同的京剧、昆曲、豫剧,实际上都建立在共通的音律理论和表演形式之上。对物质状态的本质性归类,让我们认识到所有生命分子的结构和功能,其物理根基来自于同一套量子力学规则,而不仅仅是传统化学意义上的“分子”范畴。
进一步讲,DNA、蛋白质等生物大分子之所以能够在漫长时空尺度内既稳定又兼容复杂功能,很大程度上正是由于它们属于量子固体类聚集态。聚集体内部的原子因强烈的共价键合处于稳固的“能阱”之中,这形成了高能量壁垒,为生化体系的高稳定性和高保真度提供了物理保障。
非周期性有序结构的概念
在构建大型分子聚集体时,自然界采用了两种截然不同的策略。第一种是重复性构建策略,类似于城市中的住宅楼,单位结构单元反复复制而得以大规模扩展,这就是我们熟知的晶体结构:其周期性模式一旦确立,便可无限延展而几无差异。
第二种策略则截然不同:它摒弃简单的重复,寻求个体单元高度分化和复杂功能。这正如中国古典园林的造园哲学:以苏州留园为例,园中任意一处景致,无论曲径、假山、池塘还是亭台楼阁,都是独具匠心、不可替代的;各处景观沿主题和功能有机串联,而非简单平铺堆叠。这种布局正好呼应了生命分子中信息高度有序、却非周期性重复的结构特征。
我们可以将这种结构称为“非周期性有序固体”。这个概念带来了理解生物大分子的全新视角:基因分子,乃至整个染色体体系,本质上就是一种非周期性有序固体。它们一方面拥有固体类分子的结构稳定性,一方面又能载体极其复杂的信息,这是传统周期晶体根本无法比拟的。
非周期性有序结构的真正意义,在于它能够兼容物理稳定和巨量信息两个看似矛盾的特性。一如一部鸿篇巨制的文学作品,每一字每一句既须服从整体语义和节奏,也必须精准无误地排列到位,稍有更动便失其原意。而基因分子的碱基序列或蛋白质氨基酸链,就是在这种独特结构中实现了高密度的信息编程、超高保真度的复制传递和复杂功能的动态调控。这正揭示了生命系统在根本物理层面的奥秘,连接了微观量子世界与宏观生物多样性之间的桥梁。
遗传密码的信息容量
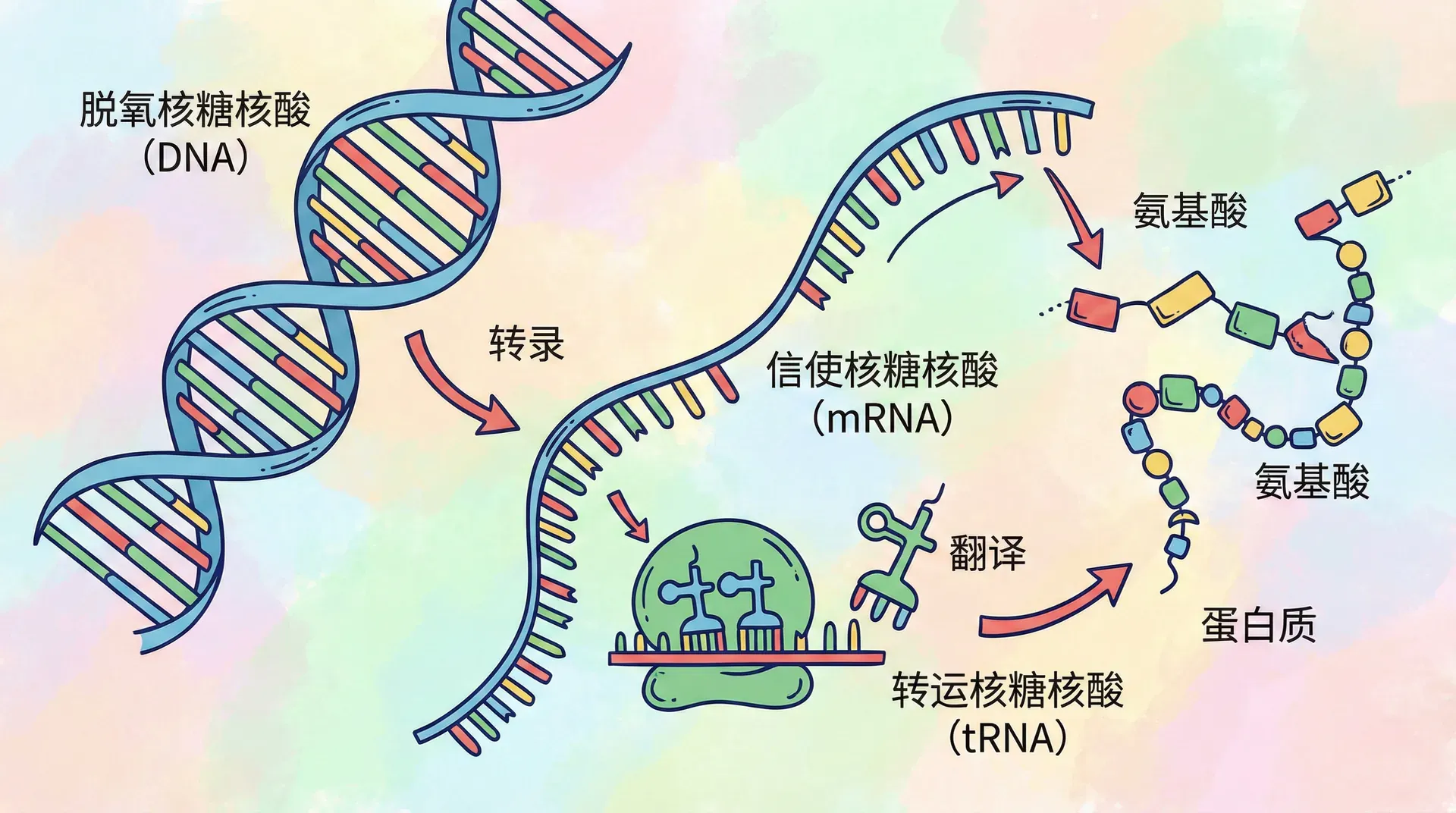
假如你用放大镜观察一粒受精卵,其细胞核几乎渺小到可以忽略不计,却蕴含着繁复生命全部的设计蓝图。这个问题长期以来令无数人疑惑:究竟如此有限的物质空间,如何能够编码和调控庞大复杂的生命系统?从今天数字科技的角度来看,这一难题甚至比芯片的数据储存还要神秘——生物的遗传物质是如何以极高密度记录下上百万的信息单元,且世世代代精准继承呢?
当我们试图理解遗传密码的高度压缩性,可以用语言和编码的例子类比:就像汉语“六书造字”,用偏旁、部首为基元,组合出上万汉字,这种结构具有显著的信息放大效应。而DNA不过由四种碱基(A、T、G、C)组成,仿佛一个仅有四个字母的“字母表”,但只要排列得足够长,信息总量便可达到天文数字。这种“指数级组合”远胜任何日常经验,哪怕只看人类基因组,每个细胞就拥有30多亿对碱基,这一串字母按照不同顺序排列出来的可能序列,已经足以容纳整个地球生物界的复杂性。
我们甚至可以更深入思考:与计算机比特“0与1”的线性堆叠不同,基因信息不仅包含碱基序列本身,还调控着基因表达、时空调度等多层级隐含编码。这就像一本小说,不仅有字面含义,还有结构、章节安排,乃至于字词搭配下的隐藏寓意。因此,遗传密码的信息容量不仅巨大,且高度多维、动态且可重组。
这种信息组合的增长规律,说明只需少量编码要素,通过排列组合,信息空间便呈爆炸式扩展。从理论到现实,这一原理早已被生命所应用,为地球上的进化和物种多样性打下了坚实基础。
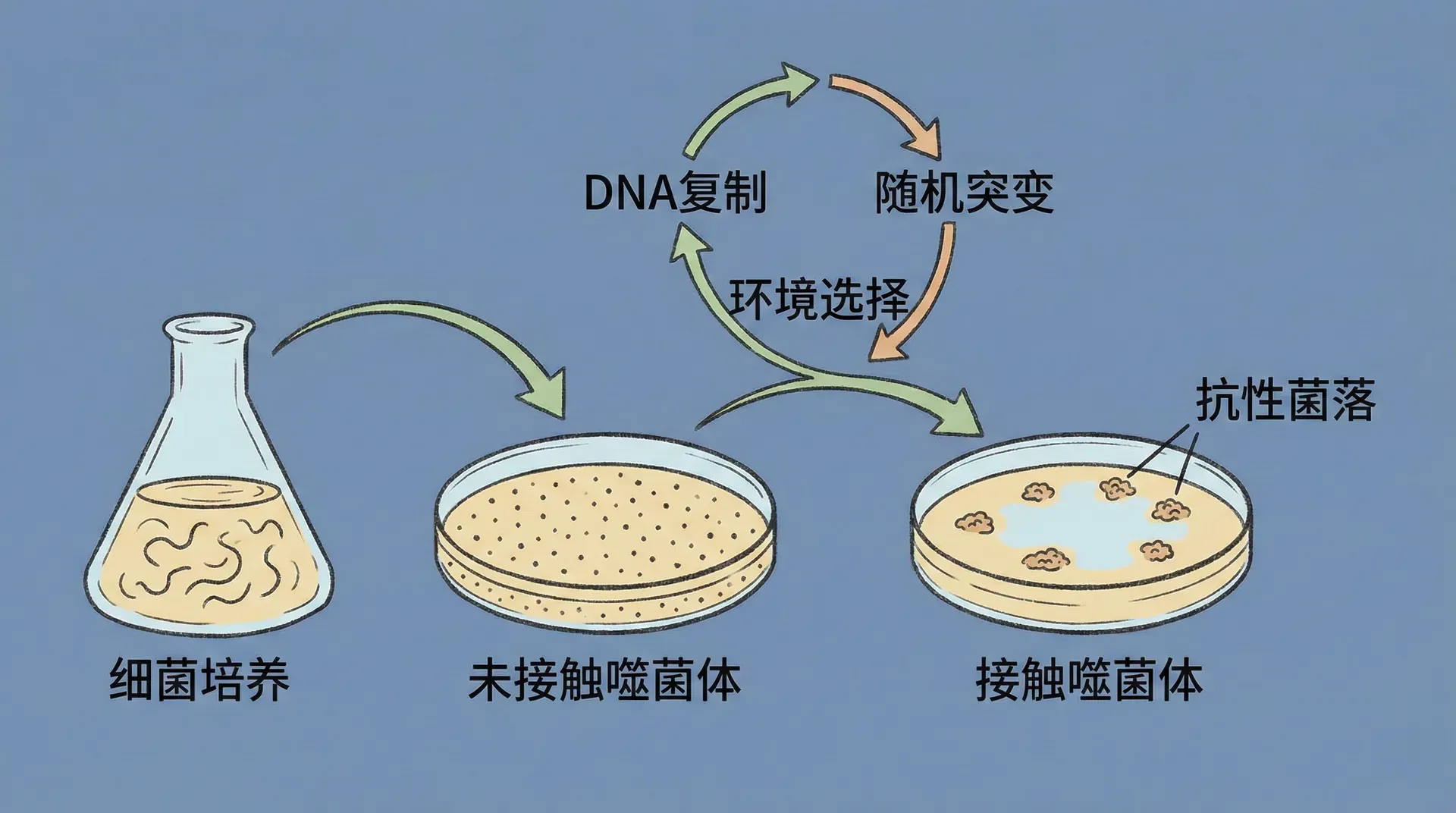
基因稳定性的实验证据
光有庞大的信息密度还不够,生命的关键还在于信息的“稳定传递”。我们不仅要存得下,还必须守得牢:基因必须经受住环境噪声、分子碰撞、化学反应等干扰,经年累月保持基础遗传信息的准确不变。这一点,在宏观世界几乎难以想象,因为多数物质和结构都很难维持千万年以上的稳定。
实验和理论在此碰头:物理模型预言,基因要做到如此高效的稳定,必须依赖极高的能量阈值。这种阈值远超日常分子的化学键能,使得自然界中随机热运动引发的自发突变概率降到极低。我们可以通过分子寿命、能垒能量的比例进行量化:
这些数字蕴含着一个道理:只有在阈值远高于平均热运动能量(kT)几十倍的情况下,分子才能拥有生物需要的超长寿命。为便于直观理解,不妨打个比方:1.8电子伏特相当于普通电池约2伏电压下,一个电子获得的能量——这对分子而言已属巨大。
自然选择与基因稳定性
有人或许会想,既然放射性、宇宙射线能诱发突变,是不是生命的自然变异主要源于这些环境因素?实际研究发现自然辐射对突变只贡献极小一部分,大多数自然突变还是源于细胞自身分子的“随机波动”。这更突显了基因自身需要设定理想的能量门槛:既不能低到轻易崩解,也不能高到一成不变,否则进化和适应便无从谈起。
在这个意义上,基因的能量阈值就是生物体自然选择的结果。回溯亿万年的自然史,无数样本被“筛选”,只有既稳又能适时突变的遗传结构才能存活优化。正如传统文化能历千年而不衰,是因为其“结构”经受了实践的反复锤炼,最终形成了系统性的优良稳定性与包容变革能力。
那些因突变而缺乏稳定性的基因往往会被“自然淘汰”,它们携带的超高突变率使后代很难长期繁衍与延续——只有能兼顾稳定与变异的基因型,才能在残酷的环境筛选中最终积累并传承下来。
温度对基因突变的影响规律
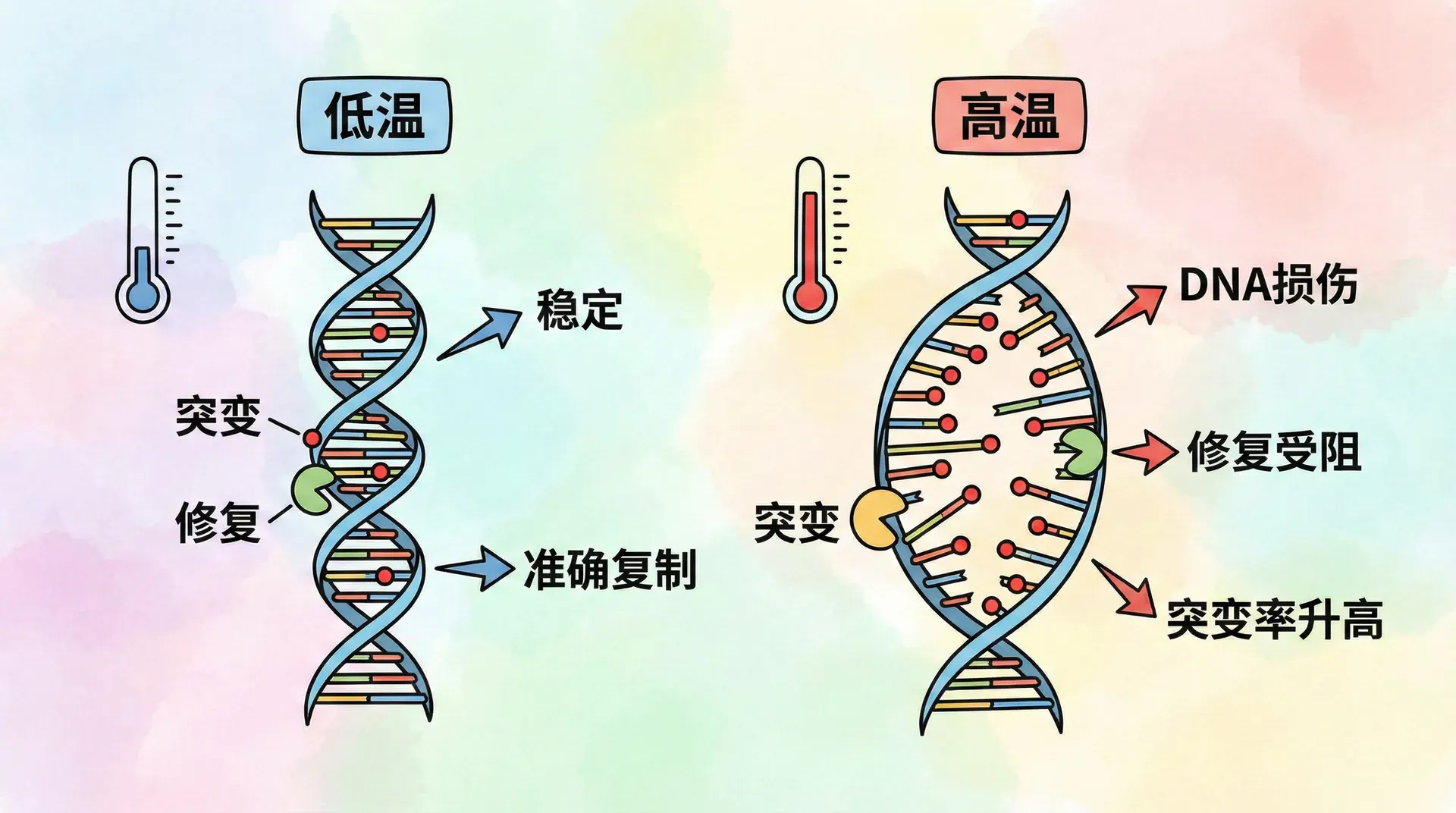
温度是影响分子运动的基础物理因子之一,那么它对基因突变的作用有多大?德尔布吕克模型的公式给出了定量答案:
这里,期望突变时间t对温度极为敏感。只需环境温度微调10℃,突变时间就会呈指数级缩短。这种精细的温度效应已通过果蝇等生物实验证实:高稳定基因的突变率会随温度上升成倍增加,而低稳定性的基因几乎不动。
我们可以把高稳定基因想象成密封极好的保险箱,需要大量外部能量才能“闯关”;而低稳定基因则像普通抽屉,即使温度升高影响微乎其微。
这些实验不仅深入检验了模型的正确性,也为我们认识更广阔的生物进化调节机制提供了量化工具。
X射线诱发突变的机制原理
通过人工手段,比如X射线辐照,人们能够明显提升基因突变率。那么X射线扮演的物理角色是什么?实验结论表明,X射线的作用类似于一种“精确打击”:它们为某一个很小的分子空间内提供大量的能量,使之瞬间越过遗传分子所需的高能阈值,从而在局部“炸开”出一次不可逆的遗传突变。
其中的机制可以这样理解——并不需要X射线本身攻击分子,而是它产生的高速电子(次级电子)在穿越分子时带入巨量能量,这股能量(常见为30电子伏特)转化为瞬时、剧烈的“局部加热”和强分子扰动,极小空间内就可促成遗传分子结构的断裂与重组。每当我们试图通过外部激发探测基因极限时,这种能量触发阈值的原理正好作为模型的重要物证之一。
模型的成功验证
将理论与实验相互印证,是科学进步的必经之路。德尔布吕克模型正是在多种独立证据(基因寿命数据、外部诱变实验、温度效应等)交叉验证中显示出极大预测力。它打破了早期对生物特异性的神秘化阐释,将生命遗传的核心机制归纳为物理与化学的基本规则之内。
我们不仅获得了对遗传稳定性“何以可能”的清晰图像,还在分子层面理解了进化与变异的源头。这个理论像一条连接自然科学多个领域的纽带,将细胞、分子、信息论和进化论如此多元的知识融于一体。
德尔布吕克模型的突破之处,在于它定量解释了生命持续性和可变性的奥妙,将看似不可调和的稳定与变异,统一到了分子物理规律之中。这为现代遗传学、分子生物学和精准医学等领域奠定了宽阔的理论根基,也指导我们以科学的视角深刻把握生命的本质。
有了这个模型,科学家们可以自信地追问:我们为什么能够遗传、为什么会突变,又为何能够在随机与秩序之间把握那样精巧的平衡。科学之河奔流不息,每一次理论的突破,都会成为未来新知识的源头活水。